Vous aimerez peut-être aussi
- Cómo se comunican las neuronas: El milagro de la transmisión sinápticaD'EverandCómo se comunican las neuronas: El milagro de la transmisión sinápticaPas encore d'évaluation
- Neurona Biologica y ArtificialDocument24 pagesNeurona Biologica y ArtificialJuanJiménezPas encore d'évaluation
- Fisiologia de Transmision de Impulso NerviosoDocument7 pagesFisiologia de Transmision de Impulso NerviosoIvantrosky MachukPas encore d'évaluation
- Redes NeuralesDocument2 pagesRedes NeuralesIsrael Lopez SaucedoPas encore d'évaluation
- Copia de Guia Sistema NerviosoDocument7 pagesCopia de Guia Sistema Nerviosojuancik7215Pas encore d'évaluation
- Trabajo Sobre La NeuronaDocument3 pagesTrabajo Sobre La Neuronacarmelo rhenals oteroPas encore d'évaluation
- Guía ProfesorDocument5 pagesGuía ProfesorDaxter Del Canto FábilaPas encore d'évaluation
- Repaso BiofisicaDocument9 pagesRepaso Biofisicamaria alejandraPas encore d'évaluation
- Final de Huamana 2Document10 pagesFinal de Huamana 2Gonzalo Nahuel LemosPas encore d'évaluation
- Fibra Nerviosa ImprimirDocument6 pagesFibra Nerviosa Imprimirerika collePas encore d'évaluation
- Impulso Nervioso - SinapsisDocument7 pagesImpulso Nervioso - SinapsisFiorellaMarPas encore d'évaluation
- 2.7 Transmisiones Sinapticas y La Unidad Motora, NeurofisiologiaDocument17 pages2.7 Transmisiones Sinapticas y La Unidad Motora, NeurofisiologiaCristobal Limones PerchezPas encore d'évaluation
- Señales Electricas de La NeuronaDocument12 pagesSeñales Electricas de La NeuronaDavid Miktlan100% (1)
- SinapsisDocument6 pagesSinapsisAna Lorena GonzalezPas encore d'évaluation
- Impulso y SinapsisDocument12 pagesImpulso y SinapsisMichelle CordovaPas encore d'évaluation
- Las NeuronasDocument5 pagesLas NeuronasLallypPas encore d'évaluation
- SensaciónDocument11 pagesSensaciónPiero PozoPas encore d'évaluation
- GuiaImpulsoNervioso - Parte1 LABDocument9 pagesGuiaImpulsoNervioso - Parte1 LABCamila AndradePas encore d'évaluation
- Conduccion Electrica en El Sistema NerviosoDocument5 pagesConduccion Electrica en El Sistema Nerviosocristhiam rojasPas encore d'évaluation
- Clase No. 07 Potencial de MembranaDocument28 pagesClase No. 07 Potencial de MembranaCamila Ñiquen ParejaPas encore d'évaluation
- SinapsisDocument6 pagesSinapsisjuan jose erasoPas encore d'évaluation
- Actividad Sistema Nervioso (1625)Document12 pagesActividad Sistema Nervioso (1625)margarita gonzalezPas encore d'évaluation
- Sistema Nervioso en AccionDocument2 pagesSistema Nervioso en AcciondianisPas encore d'évaluation
- Práctica 3. NEUROFISIOLOGÍA DEL IMPULSO NERVIOSODocument14 pagesPráctica 3. NEUROFISIOLOGÍA DEL IMPULSO NERVIOSOYosef HernándezPas encore d'évaluation
- Comunicación NeuronalDocument38 pagesComunicación NeuronalAtlamaltaPas encore d'évaluation
- Impulso Nervioso y Fibras NerviosasDocument4 pagesImpulso Nervioso y Fibras NerviosasAngelita HerreraPas encore d'évaluation
- La SinapsisDocument11 pagesLa SinapsisAnonymous f3VIazPas encore d'évaluation
- MielinizacionDocument57 pagesMielinizacionVictoria Castillo100% (1)
- Transmisión Del Impulso NerviosoDocument3 pagesTransmisión Del Impulso NerviosoAgusfa GomezPas encore d'évaluation
- Exitacion y Conduccion NeuronalDocument13 pagesExitacion y Conduccion NeuronalCésar HaroPas encore d'évaluation
- Bioquimica CerebralDocument9 pagesBioquimica CerebralStephania González DominguezPas encore d'évaluation
- Potencial de AcciónDocument5 pagesPotencial de Acciónamgraglia100% (1)
- Taller N6Document7 pagesTaller N6Grecia AlvarezPas encore d'évaluation
- Neurotransmisores y Comunicación Sináptica TallerDocument12 pagesNeurotransmisores y Comunicación Sináptica TallerNicoll Sarmiento RiberoPas encore d'évaluation
- SinapsisDocument8 pagesSinapsisloglady00Pas encore d'évaluation
- 7.c. IMPUL NERV-SINAPSIS-ACTOS NERVDocument4 pages7.c. IMPUL NERV-SINAPSIS-ACTOS NERVEster Palmira VargasPas encore d'évaluation
- Laboratorio de Impulso NerviosoDocument18 pagesLaboratorio de Impulso Nerviosomayerlis villalbaPas encore d'évaluation
- Tarea de MelisaDocument6 pagesTarea de MelisaPHONE-CENTER -CALAMARPas encore d'évaluation
- Taller Potencial de AcciónDocument3 pagesTaller Potencial de AcciónDaniellaPas encore d'évaluation
- Tema 02 Fisiologia NeuronalDocument18 pagesTema 02 Fisiologia NeuronalJonathan-Patty RiveroPas encore d'évaluation
- SinapsisDocument15 pagesSinapsisGuillermo OchoaPas encore d'évaluation
- Sinapsis TiposDocument9 pagesSinapsis TiposCastiello MnsPas encore d'évaluation
- Psicofisiologia 2Document11 pagesPsicofisiologia 2Juan Estarling TrinidadPas encore d'évaluation
- Laboratorio FisiologíaDocument6 pagesLaboratorio FisiologíaFátima Guadalupe Guevara BarahonaPas encore d'évaluation
- Procesos Que Se Dan Paso A Paso para Que Se Produzca El Impulso NerviosoDocument3 pagesProcesos Que Se Dan Paso A Paso para Que Se Produzca El Impulso NerviosoAndy LiuPas encore d'évaluation
- Marco Teórico Sistema NerviosoDocument6 pagesMarco Teórico Sistema Nerviosocamila_munozleal0% (3)
- La Neurona Unidad Funcional y EstructuralDocument68 pagesLa Neurona Unidad Funcional y Estructuralmariella104Pas encore d'évaluation
- Fisiología Del Impulso NerviosoDocument5 pagesFisiología Del Impulso NerviosoFrancisca RiveraPas encore d'évaluation
- Aplicaciones de Carga EléctricaDocument6 pagesAplicaciones de Carga EléctricaAlejandro Ü VazquezPas encore d'évaluation
- Materia: Bases BiológicasDocument8 pagesMateria: Bases BiológicasMagali CardenasPas encore d'évaluation
- Guía SinapsisDocument2 pagesGuía SinapsisMaria Eleonora Foitzick PalmaPas encore d'évaluation
- SinapsisDocument7 pagesSinapsisManuel RejonPas encore d'évaluation
- Resumen de Generación de Impulso Nerviosos y SinapsisDocument5 pagesResumen de Generación de Impulso Nerviosos y SinapsisBelenPas encore d'évaluation
- 111 Introduction To Psychology TextDocument46 pages111 Introduction To Psychology TextOliver Zapata ArmestarPas encore d'évaluation
- Investigación 1 - Aguilar Flores Juan FranciscoDocument3 pagesInvestigación 1 - Aguilar Flores Juan FranciscoFrancisco AguilarPas encore d'évaluation
- Clasificación de SinapsisDocument4 pagesClasificación de SinapsisMiguel Angel Vazquez Perez100% (1)
- Resumen CAPÍTULO 4, 5,6,7 de FerreresDocument5 pagesResumen CAPÍTULO 4, 5,6,7 de FerreresRamonPas encore d'évaluation
- 00066019Document7 pages00066019Anttonella PolancoPas encore d'évaluation
- La Conduccion Del Impulso Nervioso1Document24 pagesLa Conduccion Del Impulso Nervioso1Leidy CastroPas encore d'évaluation
- Conducción Del Impulso Nervioso Por El AxónDocument16 pagesConducción Del Impulso Nervioso Por El AxónVale LeroyerPas encore d'évaluation
- EthernetDocument9 pagesEthernetapi-3712810Pas encore d'évaluation
- Tercer ParcialDocument8 pagesTercer Parcialapi-3712810Pas encore d'évaluation
- Segundo ParcialDocument8 pagesSegundo Parcialapi-3712810Pas encore d'évaluation
- Trabajo FinalDocument7 pagesTrabajo Finalapi-3712810Pas encore d'évaluation
- Emu Lad orDocument7 pagesEmu Lad orapi-3712810Pas encore d'évaluation
- Proyecto Primer ParcialDocument8 pagesProyecto Primer Parcialapi-3712810Pas encore d'évaluation
- Generacion de Las AsDocument6 pagesGeneracion de Las Asapi-3712810Pas encore d'évaluation
- Emu Lad orDocument2 pagesEmu Lad orapi-3712810Pas encore d'évaluation
- Definicion de OffDocument1 pageDefinicion de Offapi-3712810Pas encore d'évaluation
- Cronología Del Procesador de 64 BitsDocument3 pagesCronología Del Procesador de 64 Bitsapi-3712810Pas encore d'évaluation
- Tarjetas PerforadasDocument1 pageTarjetas Perforadasapi-3712810Pas encore d'évaluation
- Clase de FayolDocument2 pagesClase de Fayolapi-3712810100% (2)
- Chipset y Arquitecturas de Microprocesadores de 64 BitsDocument3 pagesChipset y Arquitecturas de Microprocesadores de 64 Bitsapi-3720911Pas encore d'évaluation
- Generacion de Las AsDocument4 pagesGeneracion de Las Asapi-3712810Pas encore d'évaluation
- LINUXDocument4 pagesLINUXapi-3712810Pas encore d'évaluation
- Tipos de ImpresorasDocument6 pagesTipos de Impresorasapi-3712810100% (12)
- Clasificaciones de AsDocument5 pagesClasificaciones de Asapi-3712810100% (3)
- Costos Unidades EquivalentesDocument16 pagesCostos Unidades EquivalentesLeo GIPas encore d'évaluation
- Diseño de Sistemas de CosteoDocument23 pagesDiseño de Sistemas de Costeoapi-371281067% (3)
- Que Es PrologDocument6 pagesQue Es Prologapi-37128100% (1)
- Proyecto TerminadoDocument25 pagesProyecto Terminadoapi-3720911Pas encore d'évaluation
- Windows VistaDocument15 pagesWindows Vistaapi-3712810Pas encore d'évaluation
- Analisis y DiseÑoDocument1 pageAnalisis y DiseÑoapi-3712810Pas encore d'évaluation
- Primer ParcialDocument31 pagesPrimer Parcialapi-37128100% (1)
- SolicitudDocument5 pagesSolicitudapi-3712810Pas encore d'évaluation
- CaracolesDocument20 pagesCaracolesLuis Ramos OchoaPas encore d'évaluation
- Ascorbato OxidasaDocument85 pagesAscorbato OxidasaJuan RestrepoPas encore d'évaluation
- Genética MendelianaDocument17 pagesGenética Mendelianaasancheztome1550100% (4)
- Vii ConversatorioDocument2 pagesVii ConversatorioMax Bellido SanchezPas encore d'évaluation
- Tallo Cerebral o Tronco CerebralDocument7 pagesTallo Cerebral o Tronco CerebralLENARD APAZA100% (1)
- Citología CervicalDocument47 pagesCitología CervicalSalvador Luna100% (2)
- Resumen Páncreas Finn GeneserDocument3 pagesResumen Páncreas Finn GeneserLaura FMPas encore d'évaluation
- Anatomia - Tarea 4 - Maria Montes PDFDocument26 pagesAnatomia - Tarea 4 - Maria Montes PDFmariandre montes50% (2)
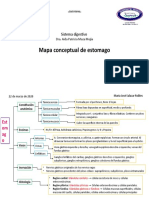
- Mapa Conceptual EstomagoDocument2 pagesMapa Conceptual EstomagoMaría José Salazar0% (1)
- Anatomía Quirúrgica Del Apéndice TextoDocument21 pagesAnatomía Quirúrgica Del Apéndice TextoEdicson MatosPas encore d'évaluation
- Clase 1-Organización Estructura y Actividad CelularDocument19 pagesClase 1-Organización Estructura y Actividad Celularljpsoft100% (3)
- Entrenamiento de La Zona Media - A. BustosDocument83 pagesEntrenamiento de La Zona Media - A. BustosLean Gabriel IturbidePas encore d'évaluation
- Ensayo de Modificación de Conducta.Document12 pagesEnsayo de Modificación de Conducta.natali100% (1)
- Cirugia OrtognaticaDocument20 pagesCirugia OrtognaticaMrArquitecto100% (1)
- Infografías Articulación de FonemasDocument18 pagesInfografías Articulación de FonemasD DPñPas encore d'évaluation
- Casos Clinicos Miembro SuperiorDocument7 pagesCasos Clinicos Miembro SuperiorJonathan RiosPas encore d'évaluation
- R.C.P. en Adultos.Document31 pagesR.C.P. en Adultos.ARTUROCUYIN100% (4)
- Electrocardiogram ADocument26 pagesElectrocardiogram AKevin Gerald Lobos MuñozPas encore d'évaluation
- Partes Del Cuerpo en NauatlDocument10 pagesPartes Del Cuerpo en NauatlMiguel Angel Vargas HernandezPas encore d'évaluation
- Competencia de Enfermeria Cancer Gastrico 2014 PDFDocument35 pagesCompetencia de Enfermeria Cancer Gastrico 2014 PDFIrvin EstebanPas encore d'évaluation
- Libro Seres ImaginariosDocument41 pagesLibro Seres Imaginariosciberleila0% (1)
- Hormonas Tiroideas (2) KDocument9 pagesHormonas Tiroideas (2) KsorayaeugeniamPas encore d'évaluation
- Actividad 4 - CuestionarioDocument8 pagesActividad 4 - CuestionarioGrinch FixPas encore d'évaluation
- TEMA 12. ZoodermatosisDocument4 pagesTEMA 12. ZoodermatosisFran VillagránPas encore d'évaluation
- Podología EquinaDocument3 pagesPodología EquinaGinebra LópezPas encore d'évaluation
- Aptos de BimlerDocument14 pagesAptos de BimlerErikaPas encore d'évaluation
- Cintura Pélvica-2Document1 pageCintura Pélvica-2Valeria FernandezPas encore d'évaluation
- NutricionDocument22 pagesNutricionHelena RomeroPas encore d'évaluation
- Danyh TrabajoDocument1 pageDanyh TrabajogladimiraPas encore d'évaluation
- Presion OsmoticaDocument6 pagesPresion OsmoticaWilson Piero Fernández VelaPas encore d'évaluation