Vous aimerez peut-être aussi
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (120)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (73)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- Anti-VEGF Use in OphthalmologyDocument231 pagesAnti-VEGF Use in OphthalmologyAnonymous 0WDdDLGg100% (1)
- Sousa 2017Document15 pagesSousa 2017PrashantPas encore d'évaluation
- (Reference Series in Biomedical Engineering) Wolfgang Holnthoner, Andrea Banfi, James Kirkpatrick, Heinz Redl (Eds.) - Vascularization For Tissue Engineering and Regenerative Medicine-Springer InternaDocument231 pages(Reference Series in Biomedical Engineering) Wolfgang Holnthoner, Andrea Banfi, James Kirkpatrick, Heinz Redl (Eds.) - Vascularization For Tissue Engineering and Regenerative Medicine-Springer InternaIsidoro OlveraPas encore d'évaluation
- Effect of Chitosajournaln JournalDocument7 pagesEffect of Chitosajournaln JournalSabrina Ceasy AnggraeniPas encore d'évaluation
- Cancer Prevention - From Mechanisms To Translational BenefitsDocument488 pagesCancer Prevention - From Mechanisms To Translational BenefitsJosé RamírezPas encore d'évaluation
- Ageing Research Reviews: A B C D e F C D eDocument18 pagesAgeing Research Reviews: A B C D e F C D eBradyPas encore d'évaluation
- Prof Rika - Konsep Dasar Biologi Sel Dan MolekularDocument33 pagesProf Rika - Konsep Dasar Biologi Sel Dan MolekularTiaradestiana AbeePas encore d'évaluation
- Effect of Long-Term Aerobic Exercise Training On Signaling Molecules in VEGF Protein Expression in Human Skeletal MuscleDocument18 pagesEffect of Long-Term Aerobic Exercise Training On Signaling Molecules in VEGF Protein Expression in Human Skeletal Muscle전유나Pas encore d'évaluation
- PI3K/AKT/mTOR Pathway in Angiogenesis: Molecular NeuroscienceDocument8 pagesPI3K/AKT/mTOR Pathway in Angiogenesis: Molecular NeuroscienceAbu KhairPas encore d'évaluation
- Cloning, Expression and Purification of Mouse Vegf (Vascular Endothelial Growth Factor) in E. Coli 01 FebDocument4 pagesCloning, Expression and Purification of Mouse Vegf (Vascular Endothelial Growth Factor) in E. Coli 01 FebGJESRPas encore d'évaluation
- Bevacizumab Product Presentation ModuleDocument35 pagesBevacizumab Product Presentation ModuleBeryl OngPas encore d'évaluation
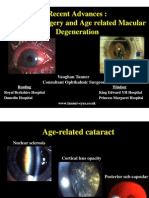
- Cataract Surgery and Age Related Macular Degeneration, Recent Advances, GP and Optometist Update 2010Document73 pagesCataract Surgery and Age Related Macular Degeneration, Recent Advances, GP and Optometist Update 2010Nyndia WeriPas encore d'évaluation
- VEGF and AngiogénesisDocument7 pagesVEGF and AngiogénesisMiranda YareliPas encore d'évaluation
- J Mol ModelingDocument12 pagesJ Mol ModelingDr. B. Venkata Raman Ph.DPas encore d'évaluation
- TranslatedDocument17 pagesTranslatedKhumaira SantaPas encore d'évaluation
- Regeneration OcularDocument40 pagesRegeneration OcularEDUARDO HUARTEPas encore d'évaluation
- Ijms 19 00311Document26 pagesIjms 19 00311Hidayat ArifinPas encore d'évaluation
- CurcuminaDocument60 pagesCurcuminaLuis Armando BuenaventuraPas encore d'évaluation
- Angiogenesis in CancerDocument4 pagesAngiogenesis in CanceranonymousPas encore d'évaluation
- Fistula ArteriovenosaDocument14 pagesFistula ArteriovenosaJoshua CajinaPas encore d'évaluation
- AngiogenezaDocument19 pagesAngiogenezaamaliaanghelPas encore d'évaluation
- Progress of Molecular Targeted Therapies For ProstatecancersDocument13 pagesProgress of Molecular Targeted Therapies For ProstatecancersNenad MladenovicPas encore d'évaluation
- Robusta Coffee On Full Thickness Wound HealingDocument6 pagesRobusta Coffee On Full Thickness Wound HealingIndah PratiwiPas encore d'évaluation
- Dafpus ProposalDocument3 pagesDafpus ProposalJantikaPas encore d'évaluation
- Molecular Mechanism of Aloe Barbadensis MillerDocument8 pagesMolecular Mechanism of Aloe Barbadensis MillerSusyana IrianiPas encore d'évaluation
- Targeting Tumor Associated Macrophages (TAMs) Via NanocarriersDocument15 pagesTargeting Tumor Associated Macrophages (TAMs) Via Nanocarriersyuvrajsingh3Pas encore d'évaluation
- Semax As A Universal Drug For Therapy and ResearchDocument12 pagesSemax As A Universal Drug For Therapy and ResearchAlvian VianPas encore d'évaluation
- Tumor AngiogenesisDocument35 pagesTumor AngiogenesisDoni Mirza KurniawanPas encore d'évaluation