Vous aimerez peut-être aussi
- Denaturing Gradient Gel Electrophoresis of β-ThalassemiaDocument2 pagesDenaturing Gradient Gel Electrophoresis of β-Thalassemiau77Pas encore d'évaluation
- ELS Amino Acid Substitutions Effects On Prot StabilityDocument8 pagesELS Amino Acid Substitutions Effects On Prot StabilityLeonardo HerreraPas encore d'évaluation
- Angela Seidl and Hans-Jurgen Hinz - The Free Energy of DNA Supercoiling Is Enthalpy-DeterminedDocument5 pagesAngela Seidl and Hans-Jurgen Hinz - The Free Energy of DNA Supercoiling Is Enthalpy-DeterminedDopamePas encore d'évaluation
- Point Mutations at Glycine-121 of Escherichia Coli Dihydrofolate Reductase: Important Roles of A Flexible Loop in The Stability and FunctionDocument8 pagesPoint Mutations at Glycine-121 of Escherichia Coli Dihydrofolate Reductase: Important Roles of A Flexible Loop in The Stability and FunctionHertzyEthanPas encore d'évaluation
- (1988) Comparison Between DNA Melting Thermodynamics and DNA Polymerase FidelityDocument5 pages(1988) Comparison Between DNA Melting Thermodynamics and DNA Polymerase Fidelitydominguezmariela465Pas encore d'évaluation
- JACS A Study of DNA Tube Formation Mechanisms Using 4 - 8 - and 12 - Helix DNA NanostructureDocument10 pagesJACS A Study of DNA Tube Formation Mechanisms Using 4 - 8 - and 12 - Helix DNA NanostructureMahirami PadronPas encore d'évaluation
- J. Patrick A. Fairclough Et Al - Chain Length Dependence of The Mean Ðeld Temperature in Poly (Oxyethylene) - Poly (Oxybutylene) Diblock CopolymersDocument3 pagesJ. Patrick A. Fairclough Et Al - Chain Length Dependence of The Mean Ðeld Temperature in Poly (Oxyethylene) - Poly (Oxybutylene) Diblock CopolymersDremHpPas encore d'évaluation
- Electrophoretic Karyotype Of: SolaniDocument3 pagesElectrophoretic Karyotype Of: SolaniVo Hoang AnhPas encore d'évaluation
- Seminario InmunoDocument5 pagesSeminario InmunoFrankly CabreraPas encore d'évaluation
- Nucleic: Efficiency E.coli by ElectroporationDocument19 pagesNucleic: Efficiency E.coli by Electroporationh_spriyaPas encore d'évaluation
- Escherichia ColiDocument10 pagesEscherichia ColisurendrasrawanPas encore d'évaluation
- Papineni 1997Document9 pagesPapineni 1997簡孟珣Pas encore d'évaluation
- Ricardo Ugarte, Carlos Bustos, Ignacio Moreno-VillosladaDocument7 pagesRicardo Ugarte, Carlos Bustos, Ignacio Moreno-VillosladaSOCKYPas encore d'évaluation
- Biopolymers - November 1982 - Marky - Calorimetric Determination of Base Stacking Enthalpies in Double Helical DNADocument10 pagesBiopolymers - November 1982 - Marky - Calorimetric Determination of Base Stacking Enthalpies in Double Helical DNAhajardreamhighPas encore d'évaluation
- Melting Behavior of Pseudoknots Containing Adjacent GC and AT Rich DomainsDocument7 pagesMelting Behavior of Pseudoknots Containing Adjacent GC and AT Rich DomainsMiftahul KautsariPas encore d'évaluation
- Bi 00444 A 006Document8 pagesBi 00444 A 006Moh SyaifudinPas encore d'évaluation
- Jurnal TermalDocument6 pagesJurnal TermalZulfi RahmanPas encore d'évaluation
- PhysRevC 65 014308Document10 pagesPhysRevC 65 014308Ravi Sankar Babu BalabhadrapatruniPas encore d'évaluation
- 249 2012 Article 865Document12 pages249 2012 Article 865Ivan VelascoPas encore d'évaluation
- Anja Dietrich2002Document21 pagesAnja Dietrich2002OhisverPas encore d'évaluation
- 0532 HTMLDocument6 pages0532 HTMLyuPas encore d'évaluation
- Force Induced DNA Melting: (Prepared For The Special Issue of JPCM On "DNA Melting", Edited by Ralf Blossey)Document23 pagesForce Induced DNA Melting: (Prepared For The Special Issue of JPCM On "DNA Melting", Edited by Ralf Blossey)Soma GhoshPas encore d'évaluation
- FAD Used As A Mediator in The Electron Transfer Between Platinum and Several BiomoleculesDocument11 pagesFAD Used As A Mediator in The Electron Transfer Between Platinum and Several BiomoleculesLata DeshmukhPas encore d'évaluation
- Comparative Analysis of Gentamicin Resistance in Staphylococcus AureusDocument3 pagesComparative Analysis of Gentamicin Resistance in Staphylococcus Aureusyanita rahmadhaniPas encore d'évaluation
- Is Tetramethylene An Intermediate?: Figure 1. Contour Plot A DisDocument2 pagesIs Tetramethylene An Intermediate?: Figure 1. Contour Plot A DisJosé CortésPas encore d'évaluation
- On The Truncation of Long-Range Electrostatic Interactions in DNADocument17 pagesOn The Truncation of Long-Range Electrostatic Interactions in DNADavidFurmanPas encore d'évaluation
- Department of Medical Enzymology, Laboratory of Biochemistry, University of Amsterdam, Amsterdam (The Netherlands)Document9 pagesDepartment of Medical Enzymology, Laboratory of Biochemistry, University of Amsterdam, Amsterdam (The Netherlands)RiridJatmikoPas encore d'évaluation
- Acs 2013Document1 pageAcs 2013api-255627078Pas encore d'évaluation
- GIAO/DFT Evaluation of C NMR Chemical Shifts of Selected Acetals Based On DFT Optimized GeometriesDocument8 pagesGIAO/DFT Evaluation of C NMR Chemical Shifts of Selected Acetals Based On DFT Optimized Geometriesganesh_orcrdPas encore d'évaluation
- O. Varnavski, G. Menkir, T. Goodson III and P. L. Burn - Ultrafast Polarized Fluorescence Dynamics in An Organic DendrimerDocument3 pagesO. Varnavski, G. Menkir, T. Goodson III and P. L. Burn - Ultrafast Polarized Fluorescence Dynamics in An Organic DendrimerGlade680Pas encore d'évaluation
- 1) Electrophoresis - IntroductionDocument12 pages1) Electrophoresis - IntroductionMuhammad AdeelPas encore d'évaluation
- Mutagenesis High-Affinity: Site-Directed Streptavidin-Biotin Complex: Contributions Tryptophan Residues 79, 108Document5 pagesMutagenesis High-Affinity: Site-Directed Streptavidin-Biotin Complex: Contributions Tryptophan Residues 79, 108Guhan KAPas encore d'évaluation
- Electronic Supplementary Information (ESI) : 2. ExperimentalDocument5 pagesElectronic Supplementary Information (ESI) : 2. ExperimentalEsperanza SAPas encore d'évaluation
- Kumar Nussinov Biochem 2001 Mesophile Thermophile StabilityDocument14 pagesKumar Nussinov Biochem 2001 Mesophile Thermophile StabilityJames NanorPas encore d'évaluation
- Maxam and Gilbert DNA SequencingDocument5 pagesMaxam and Gilbert DNA SequencingnipanshuPas encore d'évaluation
- The Polymerase Chain Reaction and Sequence-Tagged SitesDocument7 pagesThe Polymerase Chain Reaction and Sequence-Tagged SitesPisi MicaPas encore d'évaluation
- 1 s2.0 0032386183900514 MainDocument7 pages1 s2.0 0032386183900514 MainGeorge KafentzisPas encore d'évaluation
- Kang 2005Document4 pagesKang 2005Quan LinhPas encore d'évaluation
- Reaccion de Eliminacion E y ZDocument5 pagesReaccion de Eliminacion E y ZKarelis GutierrezPas encore d'évaluation
- A DFT-Based Computational Study On Hydrogen Tunneling Phenomenon in The Isomerization of MethylhydroxycarbeneDocument10 pagesA DFT-Based Computational Study On Hydrogen Tunneling Phenomenon in The Isomerization of MethylhydroxycarbenejumaidilawalPas encore d'évaluation
- Hydrated and Dehydrated Tertiary Interactions-Opening and Closing-Of A Four-Helix Bundle PeptideDocument9 pagesHydrated and Dehydrated Tertiary Interactions-Opening and Closing-Of A Four-Helix Bundle Peptide666667Pas encore d'évaluation
- Geometry Changes of A Cu (I) Phenanthroline Complex On Photoexcitation in A Confining Medium by Time-Resolved X-Ray DiffractionDocument2 pagesGeometry Changes of A Cu (I) Phenanthroline Complex On Photoexcitation in A Confining Medium by Time-Resolved X-Ray DiffractionJoakin BahamondesPas encore d'évaluation
- Application of Van't Hoff Eqn 2Document5 pagesApplication of Van't Hoff Eqn 2Kanan PatelPas encore d'évaluation
- Gupta Spin Coherence in Semiconductor Quantum DotsDocument4 pagesGupta Spin Coherence in Semiconductor Quantum DotsEddy YusufPas encore d'évaluation
- Eco Ri y Otras ReaccionesDocument13 pagesEco Ri y Otras ReaccionesGustavo CgPas encore d'évaluation
- LSM1102 - Studies On Transformation of Escherichia Coli With PlasmidsDocument24 pagesLSM1102 - Studies On Transformation of Escherichia Coli With Plasmidsgivena2ndchancePas encore d'évaluation
- Molecular Insights Into The Gel To Liquid-Crystalline Phase Transition of Phospholipid BilayersDocument1 pageMolecular Insights Into The Gel To Liquid-Crystalline Phase Transition of Phospholipid BilayerslwnnrcPas encore d'évaluation
- Defect States in Perovskite Solar Cells Associated With Hysteresis and PerformanceDocument5 pagesDefect States in Perovskite Solar Cells Associated With Hysteresis and PerformanceNILOFARPas encore d'évaluation
- 89 JBC - 264 - 17309Document7 pages89 JBC - 264 - 17309Gregory BurrowsPas encore d'évaluation
- Pi Is 0092867409014342Document17 pagesPi Is 0092867409014342Roger TsengPas encore d'évaluation
- A Solid State NMR Study of Dynamics in A Hydrated Salivary Peptide Adsorbed To HydroxyapatiteDocument2 pagesA Solid State NMR Study of Dynamics in A Hydrated Salivary Peptide Adsorbed To HydroxyapatiteSveti JeronimPas encore d'évaluation
- Workshop 1Document13 pagesWorkshop 1DaisyPas encore d'évaluation
- Molecular Systems Biology Assignment 2: Thus It Represents The Weighted Harmonic Mean ofDocument3 pagesMolecular Systems Biology Assignment 2: Thus It Represents The Weighted Harmonic Mean ofArjun KumarPas encore d'évaluation
- Denaturing High Performance Liquid ChromatographyDocument3 pagesDenaturing High Performance Liquid ChromatographyRONAK LASHKARIPas encore d'évaluation
- Ref 6Document4 pagesRef 6fathmaPas encore d'évaluation
- Vrouw, Mar 2011Document4 pagesVrouw, Mar 2011emediagePas encore d'évaluation
- Original PaperDocument7 pagesOriginal PaperElbanhawy4EverPas encore d'évaluation
- Ojocollagen FTIR Payne1988Document12 pagesOjocollagen FTIR Payne1988FranciscoPas encore d'évaluation
- Denaturasi DnaDocument8 pagesDenaturasi DnadetiPas encore d'évaluation
- Reviews in Computational Chemistry, Volume 31D'EverandReviews in Computational Chemistry, Volume 31Abby L. ParrillPas encore d'évaluation
- Presentation For Gender and Society 2Document24 pagesPresentation For Gender and Society 2Fhebelyn TaborPas encore d'évaluation
- Science 8 Q4M1 Digestive System 2021 2022Document5 pagesScience 8 Q4M1 Digestive System 2021 2022carl helloPas encore d'évaluation
- Testicular CancerDocument5 pagesTesticular CancerypintohPas encore d'évaluation
- Module 4 Digestive SystemDocument9 pagesModule 4 Digestive SystemupadhiPas encore d'évaluation
- Report of The Experimental Results of Drossinakis IAWG Korotkov 2012Document21 pagesReport of The Experimental Results of Drossinakis IAWG Korotkov 2012magi44kenn100% (1)
- Diagnosis and Treatment of Obstructive Sleep Apnea in Adults PDFDocument7 pagesDiagnosis and Treatment of Obstructive Sleep Apnea in Adults PDFkartini ciatawiPas encore d'évaluation
- Abbot, Diabetes ManagementDocument1 pageAbbot, Diabetes ManagementMelissa Grace DizonPas encore d'évaluation
- Total Carbohydrate Colorimetric Assay KitDocument2 pagesTotal Carbohydrate Colorimetric Assay Kitwoomh94Pas encore d'évaluation
- Cell Organelle Chart Key 2011Document3 pagesCell Organelle Chart Key 2011Angel Silva Vicente50% (2)
- And Um-101Document3 pagesAnd Um-101ver_at_workPas encore d'évaluation
- Rula SheetDocument2 pagesRula SheetCarl Christian CabelloPas encore d'évaluation
- SciaticaDocument3 pagesSciaticaNick BantoloPas encore d'évaluation
- Signal TransductionDocument38 pagesSignal TransductionIndraPramanaIdaBagusPas encore d'évaluation
- CBC Reviewer Anaphy LabDocument9 pagesCBC Reviewer Anaphy LabARVINE JUSTINE CORPUZPas encore d'évaluation
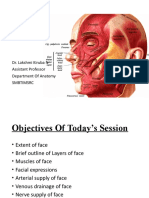
- Dr. Lakshmi Kiruba N Assistant Professor Department of Anatomy SmbtimsrcDocument25 pagesDr. Lakshmi Kiruba N Assistant Professor Department of Anatomy Smbtimsrclakshmi kirubaPas encore d'évaluation
- Drosophila Melanogaster Models of Motor Neuron Disease 2013 (Edited by Ruben Cauchi)Document239 pagesDrosophila Melanogaster Models of Motor Neuron Disease 2013 (Edited by Ruben Cauchi)Dr.Gill_Grissom100% (1)
- CropSci101 Module 3Document11 pagesCropSci101 Module 3Jetro Neil GapasinPas encore d'évaluation
- Acute Miocard InfarkDocument32 pagesAcute Miocard InfarkhansPas encore d'évaluation
- Electromyography I PDFDocument13 pagesElectromyography I PDFkeithtrahPas encore d'évaluation
- Cardiac Embryology 1Document18 pagesCardiac Embryology 1Natalia IhalauwPas encore d'évaluation
- 24 Cerebral Blood FlowDocument31 pages24 Cerebral Blood FlowZuhaib Ahmed100% (1)
- Medscape Imperforate AnusDocument22 pagesMedscape Imperforate AnusVonny RiskaPas encore d'évaluation
- Data Alkes 2020 TerbaruDocument23 pagesData Alkes 2020 TerbaruRoyliPas encore d'évaluation
- (Jagaran Josh) Science PDFDocument136 pages(Jagaran Josh) Science PDFPiyush KumarPas encore d'évaluation
- Bioreaction Engineering Principles Nielsen Villadsen PDFDocument554 pagesBioreaction Engineering Principles Nielsen Villadsen PDFMuchachos del Rio50% (2)
- CVD PDFDocument135 pagesCVD PDFselflessdoctorPas encore d'évaluation
- Kidney Structure (14.1.3) EdExcel A Level Biology Revision Notes 2022Document2 pagesKidney Structure (14.1.3) EdExcel A Level Biology Revision Notes 2022Steven FieldsPas encore d'évaluation
- How Cells Harvest Chemical EnergyDocument75 pagesHow Cells Harvest Chemical EnergylinPas encore d'évaluation
- Reviewer 18 4 BPPK MCQDocument5 pagesReviewer 18 4 BPPK MCQBio Data50% (2)
- Steroids: Understanding Causes, Effects, and InformationDocument9 pagesSteroids: Understanding Causes, Effects, and InformationChris weberPas encore d'évaluation