Vous aimerez peut-être aussi
- Reconstructing Carotenoid-Based and Structural Coloration in Fossil Skin, McNamara Et Al., 2016Document9 pagesReconstructing Carotenoid-Based and Structural Coloration in Fossil Skin, McNamara Et Al., 2016planessPas encore d'évaluation
- Profesorul Virgil Ghiurcă (1927-2008), Promotor Al Gemologiei În RomâniaDocument14 pagesProfesorul Virgil Ghiurcă (1927-2008), Promotor Al Gemologiei În RomâniaplanessPas encore d'évaluation
- Fossil Scales Illuminate The Early Evolution of Lepidopterans and Structural Colors, Zhang Et Al., 2018Document8 pagesFossil Scales Illuminate The Early Evolution of Lepidopterans and Structural Colors, Zhang Et Al., 2018planessPas encore d'évaluation
- Lithostratigraphy of Cyclic Continental To Marine Eocene Deposits in NW Transylvania, Romania, Popescu, 1984 PDFDocument24 pagesLithostratigraphy of Cyclic Continental To Marine Eocene Deposits in NW Transylvania, Romania, Popescu, 1984 PDFplanessPas encore d'évaluation
- Eocene From Bosna and Hercegovina, Soklic, 2001Document604 pagesEocene From Bosna and Hercegovina, Soklic, 2001planess100% (1)
- Eocene Sea Cows (Sirenia, Mammalia) From Hungary, Kordos, 2002Document6 pagesEocene Sea Cows (Sirenia, Mammalia) From Hungary, Kordos, 2002planessPas encore d'évaluation
- Maastrichtian Dinosaurs in SW Transylvania (Romania), Codrea Et Al., 2010Document7 pagesMaastrichtian Dinosaurs in SW Transylvania (Romania), Codrea Et Al., 2010planessPas encore d'évaluation
- Biostratigraphy of The Bartonian Deposits From Gilău Area (NW Transylvania, Romania)Document14 pagesBiostratigraphy of The Bartonian Deposits From Gilău Area (NW Transylvania, Romania)planessPas encore d'évaluation
- Formatiunile Mezozoice Din Sud-Vestul Bazinului Rusca Montană Bucur - Ea - 1985Document27 pagesFormatiunile Mezozoice Din Sud-Vestul Bazinului Rusca Montană Bucur - Ea - 1985planessPas encore d'évaluation
- VERTEBRATE PRESERVATION IN FLUVIAL CHANNELS - Behrensmeyer 1988 PDFDocument17 pagesVERTEBRATE PRESERVATION IN FLUVIAL CHANNELS - Behrensmeyer 1988 PDFplanessPas encore d'évaluation
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5795)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (400)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (74)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Early Complications of MIDocument11 pagesEarly Complications of MIjen262004Pas encore d'évaluation
- Bowel Nosodes Power PointDocument15 pagesBowel Nosodes Power PointTheresa Dale83% (6)
- Aplastic Anemia Case Study FINALDocument53 pagesAplastic Anemia Case Study FINALMatthew Rich Laguitao Cortes100% (6)
- Chapter 24)Document10 pagesChapter 24)Hazel LopezPas encore d'évaluation
- Management Organizational Behavior: Individuals in Organizations: Perception, Personality, and Cultural DifferencesDocument37 pagesManagement Organizational Behavior: Individuals in Organizations: Perception, Personality, and Cultural DifferencestesfalemPas encore d'évaluation
- Extended SexDocument18 pagesExtended SexMaría Alejandra MartínezPas encore d'évaluation
- Inhaled Anesthesia For BirdsDocument18 pagesInhaled Anesthesia For BirdsBianca PaludetoPas encore d'évaluation
- Format Ketikan LabDocument9 pagesFormat Ketikan Labapril babyPas encore d'évaluation
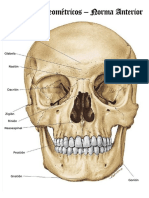
- Puntos Craneometricos CompressDocument13 pagesPuntos Craneometricos CompressLaura PPas encore d'évaluation
- Lecture 11. Avian Nervous SystemDocument6 pagesLecture 11. Avian Nervous SystemCube MaximPas encore d'évaluation
- The Peripheral Nervous System Branches Throughout The Body Worksheet1Document2 pagesThe Peripheral Nervous System Branches Throughout The Body Worksheet1Shekeyva SmalleyPas encore d'évaluation
- List of Mineral Function and DeficiencyDocument16 pagesList of Mineral Function and Deficiencyketavmodi100% (8)
- Drug FeaturesDocument9 pagesDrug FeaturesLeah Claudia de OcampoPas encore d'évaluation
- Fluids and Transport: Fundamentals of Anatomy & PhysiologyDocument161 pagesFluids and Transport: Fundamentals of Anatomy & Physiologyann aquinoPas encore d'évaluation
- 22 Stretches To Improve Your Flexibility For PoleDocument10 pages22 Stretches To Improve Your Flexibility For PoleSarah86% (7)
- The Concise Human Body Book An Illustrated Guide To Its Structure, Function, and Disorders PDFDocument322 pagesThe Concise Human Body Book An Illustrated Guide To Its Structure, Function, and Disorders PDFBriyith Triana Rondon100% (4)
- Neurexan Patient Flyer ANG May 2010 7311Document2 pagesNeurexan Patient Flyer ANG May 2010 7311Waqar GhoryPas encore d'évaluation
- And Um-101Document3 pagesAnd Um-101ver_at_workPas encore d'évaluation
- Zabrina Joana DDocument4 pagesZabrina Joana DEmman BalidoPas encore d'évaluation
- Anatomy and Physiology of NeuronsDocument4 pagesAnatomy and Physiology of NeuronsVan Jasper Pimentel BautistaPas encore d'évaluation
- PP03L039 - Disorders of The PancreasDocument41 pagesPP03L039 - Disorders of The Pancreasapi-3805855100% (3)
- Marieb ch3bDocument25 pagesMarieb ch3bapi-229554503100% (1)
- Apex o GenesisDocument54 pagesApex o GenesisbrahmannaPas encore d'évaluation
- 12 Biology Notes Ch06 Molecular Basis of InheritanceDocument6 pages12 Biology Notes Ch06 Molecular Basis of InheritanceInderpal SinghPas encore d'évaluation
- GastritisDocument10 pagesGastritisMario100% (2)
- Chapter 20 - Embryology of The Genitourinary Tract - Campbell - Walsh-Wein UROLOGY 12thDocument40 pagesChapter 20 - Embryology of The Genitourinary Tract - Campbell - Walsh-Wein UROLOGY 12thkrisnaPas encore d'évaluation
- Iggy Med Surg Test Bank Chapter 005Document19 pagesIggy Med Surg Test Bank Chapter 005Tracy Bartell80% (10)
- Progr. PR., Simpozion Boli Genet Metab, Cluj-Napoca, 7-8 IunieDocument3 pagesProgr. PR., Simpozion Boli Genet Metab, Cluj-Napoca, 7-8 IunieDoina Ababii UrsanPas encore d'évaluation
- Semi-Detailed Lesson Plan in SCIENCE Grade 9Document4 pagesSemi-Detailed Lesson Plan in SCIENCE Grade 9Sheryll Cayme SalindoPas encore d'évaluation
- Vital SignsDocument3 pagesVital SignsNicole NipasPas encore d'évaluation