Vous aimerez peut-être aussi
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- 2ND Activity Frezel EjemparDocument5 pages2ND Activity Frezel EjemparRONALD ARTILLEROPas encore d'évaluation
- Understanding Fat OxidationDocument10 pagesUnderstanding Fat OxidationPaulo CesarPas encore d'évaluation
- Eating DHEADocument18 pagesEating DHEAYutt PlermpiromPas encore d'évaluation
- Congenital HypothyroidismDocument21 pagesCongenital HypothyroidismIkhsan Nurkholis SudradjatPas encore d'évaluation
- Glia Accumulate Evidence That Actions Are Futile and Suppress Unsuccessful BehaviorDocument37 pagesGlia Accumulate Evidence That Actions Are Futile and Suppress Unsuccessful Behaviornabru cordeiroPas encore d'évaluation
- CXC CSEC HSB January 2017 P2 PDFDocument17 pagesCXC CSEC HSB January 2017 P2 PDFSayyid Muhammad Aqeed AabidiPas encore d'évaluation
- Brain Gym ExercisesDocument9 pagesBrain Gym ExercisesMuza RasidiPas encore d'évaluation
- Unit 4 Introduction To Bones and The SkullDocument22 pagesUnit 4 Introduction To Bones and The SkullrynPas encore d'évaluation
- Biology 209 Lab ManualDocument128 pagesBiology 209 Lab ManualikomalPas encore d'évaluation
- Nausea and VomitingDocument5 pagesNausea and VomitingIndra Setya PermanaPas encore d'évaluation
- Epistasis PracticeDocument1 pageEpistasis PracticeChristian MirandaPas encore d'évaluation
- San Lorenzo Ruiz College of Ormoc, Inc. College of Nursing MC1-Lab Anatomy & Physiology Learning & Performance Task 1Document4 pagesSan Lorenzo Ruiz College of Ormoc, Inc. College of Nursing MC1-Lab Anatomy & Physiology Learning & Performance Task 1MAVIE GOMEZPas encore d'évaluation
- Nursing Care PlanDocument22 pagesNursing Care PlanjamPas encore d'évaluation
- Beta Oxidation of FattyacidDocument26 pagesBeta Oxidation of FattyacidLipsa Rani Panda100% (1)
- Holter SolutionsDocument4 pagesHolter SolutionsgimenPas encore d'évaluation
- Airway: Nursing Care PlanDocument3 pagesAirway: Nursing Care PlanPhoebe Kyles CammaPas encore d'évaluation
- Bio 130 Chapter 4 NotesDocument12 pagesBio 130 Chapter 4 Notesgilissa100% (1)
- February 2021 Teaching Roster - MBBSDocument4 pagesFebruary 2021 Teaching Roster - MBBSasjxhiBBniu01j2Pas encore d'évaluation
- MultraumaDocument9 pagesMultraumaDaniel Andres ForeroPas encore d'évaluation
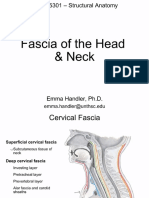
- Fascia of The Head & NeckDocument8 pagesFascia of The Head & NeckMahesh Jung KhadkaPas encore d'évaluation
- In Focus Periodic Paralysis PDFDocument8 pagesIn Focus Periodic Paralysis PDFLisa Pratiwi RPas encore d'évaluation
- Anesthesia in Hepatic DiseaseDocument3 pagesAnesthesia in Hepatic DiseaseRoxana SurliuPas encore d'évaluation
- Science 9th Chapter 5 Cell Biology NotesDocument4 pagesScience 9th Chapter 5 Cell Biology NotesavanishPas encore d'évaluation
- 11 Biology Notes ch18 Body Fluids and Circulation PDFDocument8 pages11 Biology Notes ch18 Body Fluids and Circulation PDFSumit SinghalPas encore d'évaluation
- Biology Chapter 3 Notes PDFDocument5 pagesBiology Chapter 3 Notes PDFLisa KylPas encore d'évaluation
- Cells TestDocument3 pagesCells TestSabita SinghPas encore d'évaluation
- Information For You: An Ectopic PregnancyDocument25 pagesInformation For You: An Ectopic PregnancyAnonymous layszXmsPas encore d'évaluation
- Small Animal Clinical NutritionDocument57 pagesSmall Animal Clinical NutritionJairo Pereira100% (1)
- A Ciência Da Esgrima: Implicações Da Performance e Prevenção de LesõesDocument18 pagesA Ciência Da Esgrima: Implicações Da Performance e Prevenção de LesõesDaniel GonçalvesPas encore d'évaluation
- Reseptor Kanal Ion Target ObatDocument65 pagesReseptor Kanal Ion Target ObatFitry Ana UtamPas encore d'évaluation