Vous aimerez peut-être aussi
- Saf Frey 2013Document14 pagesSaf Frey 2013Leonardo AzevedoPas encore d'évaluation
- Editorial: James L. Oschman, PHDDocument4 pagesEditorial: James L. Oschman, PHDKaycsa AdrianaPas encore d'évaluation
- Editorial: James L. Oschman, PHDDocument5 pagesEditorial: James L. Oschman, PHDSofia marisa fernandesPas encore d'évaluation
- 1disfagia en El AncianoDocument13 pages1disfagia en El AncianonadiabizesepulvedaPas encore d'évaluation
- Fmed 10 1335119Document3 pagesFmed 10 1335119dr.martynchukPas encore d'évaluation
- Cellular Changes in The ENSDocument12 pagesCellular Changes in The ENSErnesto Ochoa MonroyPas encore d'évaluation
- Theories of AgingDocument12 pagesTheories of AgingBulea Paula100% (2)
- The Science of LongevityDocument5 pagesThe Science of LongevityTogrul MusabeyliPas encore d'évaluation
- A Review of Swallow Timing in The ElderlyDocument15 pagesA Review of Swallow Timing in The ElderlyMarian J. NietoPas encore d'évaluation
- NutritionDocument3 pagesNutritionEdith Omoze Arughu-JosephPas encore d'évaluation
- Translational Endocrinology of Bone: Reproduction, Metabolism, and the Central Nervous SystemD'EverandTranslational Endocrinology of Bone: Reproduction, Metabolism, and the Central Nervous SystemGerard KarsentyPas encore d'évaluation
- Editorial: James L. Oschman, PHDDocument5 pagesEditorial: James L. Oschman, PHDTorentSpyPas encore d'évaluation
- Ajpendo 00295 2016Document12 pagesAjpendo 00295 2016Nidhi SharmaPas encore d'évaluation
- The Science of Aging How Our Bodies Change as We Get OlderD'EverandThe Science of Aging How Our Bodies Change as We Get OlderPas encore d'évaluation
- The Chronobiology, Etiology and Pathophysiology of ObesityDocument2 pagesThe Chronobiology, Etiology and Pathophysiology of ObesityDIANPas encore d'évaluation
- Lifestyle Modifications and Erectile Dysfunction: What Can Be Expected?Document6 pagesLifestyle Modifications and Erectile Dysfunction: What Can Be Expected?APas encore d'évaluation
- Research Paper PhysiologyDocument7 pagesResearch Paper Physiologym0d1p1fuwub2100% (1)
- 1 s2.0 S0092867415001865 Main PDFDocument13 pages1 s2.0 S0092867415001865 Main PDFtlidiaPas encore d'évaluation
- 10.1007-S00210-008-027serotonin Pharmacology in The Gastrointestinal Tract: A ReviewDocument23 pages10.1007-S00210-008-027serotonin Pharmacology in The Gastrointestinal Tract: A ReviewanataeusPas encore d'évaluation
- 1 s2.0 S0531556523000992 MainDocument10 pages1 s2.0 S0531556523000992 MainNatalia BeltranPas encore d'évaluation
- RRL-WPS OfficeDocument14 pagesRRL-WPS Officeprincelaureto2Pas encore d'évaluation
- Boehringer Simp Obst Kronik Ukrida 070413 (Chudahman Manan)Document47 pagesBoehringer Simp Obst Kronik Ukrida 070413 (Chudahman Manan)strawberry piePas encore d'évaluation
- Sleep and Circadian Rhythms: Key Components in The Regulation of Energy MetabolismDocument10 pagesSleep and Circadian Rhythms: Key Components in The Regulation of Energy MetabolismAacg MeryendPas encore d'évaluation
- Dysphagia in The Elderly: Ianessa A. Humbert, PHD, Joanne Robbins, PHDDocument14 pagesDysphagia in The Elderly: Ianessa A. Humbert, PHD, Joanne Robbins, PHDVallery CamilaPas encore d'évaluation
- Acupuncture-Based ModalitiesDocument9 pagesAcupuncture-Based ModalitiesFelipe MaiaPas encore d'évaluation
- Lack of Regular Physical Exercise or Too Much Inactivity: John P. Thyfault and Frank W. BoothDocument5 pagesLack of Regular Physical Exercise or Too Much Inactivity: John P. Thyfault and Frank W. BoothJor ChavezPas encore d'évaluation
- FLOR Subclinical Hypothyroidism (SCH) Is Not Associated With Metabolic DerangementDocument6 pagesFLOR Subclinical Hypothyroidism (SCH) Is Not Associated With Metabolic DerangementPeter GalvezPas encore d'évaluation
- Nutrition, Physical Activity, and Quality of Life in Older AdultsDocument6 pagesNutrition, Physical Activity, and Quality of Life in Older AdultsDanusa AlvesPas encore d'évaluation
- Aging Adiposity and Calorie Restriction Fontana2007Document9 pagesAging Adiposity and Calorie Restriction Fontana2007rhabarberrhabarberPas encore d'évaluation
- Senam ErgonomikDocument8 pagesSenam ErgonomikAngeliaPas encore d'évaluation
- Senam ErgonomicDocument8 pagesSenam ErgonomicBella Desy Vita AuliyaPas encore d'évaluation
- Sex-Dependent Metabolic, Neuroendocrine, and Cognitive Responses To Dietary Energy Restriction and ExcessDocument16 pagesSex-Dependent Metabolic, Neuroendocrine, and Cognitive Responses To Dietary Energy Restriction and ExcessBillard StéphanePas encore d'évaluation
- Forever Young: Health & Spiritual SeriesD'EverandForever Young: Health & Spiritual SeriesÉvaluation : 5 sur 5 étoiles5/5 (1)
- Best Practice & Research Clinical Endocrinology & MetabolismDocument16 pagesBest Practice & Research Clinical Endocrinology & MetabolismvipereejayPas encore d'évaluation
- Challenging the Limits of the Human Life Span: - Can We Live Longer Than 120 Years - New GuidelinesD'EverandChallenging the Limits of the Human Life Span: - Can We Live Longer Than 120 Years - New GuidelinesPas encore d'évaluation
- Undulating Changes in Human Plasma Proteome Profiles Across The LifespanDocument22 pagesUndulating Changes in Human Plasma Proteome Profiles Across The LifespanЗлойБобрPas encore d'évaluation
- Comps Paper NotesDocument17 pagesComps Paper NotesMarcus SuvantoPas encore d'évaluation
- The Aging of Skeletal Muscle and Potential Therapeutic Effects of Extracts From Edible and Inedible PlantsDocument19 pagesThe Aging of Skeletal Muscle and Potential Therapeutic Effects of Extracts From Edible and Inedible Plantsaaysha guptaPas encore d'évaluation
- Geriatric PharmacologyDocument17 pagesGeriatric PharmacologyLuciana OliveiraPas encore d'évaluation
- Gastrointestinal Physiology: June 2018Document18 pagesGastrointestinal Physiology: June 2018Bruno MarquesPas encore d'évaluation
- JournalDocument1 pageJournalJEVEE JEAN BETTERPas encore d'évaluation
- A. Biology of Aging PDFDocument9 pagesA. Biology of Aging PDFlolipopckpPas encore d'évaluation
- Gastrointestinal Physiology, JohnsonDocument310 pagesGastrointestinal Physiology, JohnsonSubhajit Chakraborty80% (5)
- Effect of Intermittent Fasting On Brain Neurotransmitters, Neutrophils Phagocytic Activity, and Histopathological Finding in Some Organs in RatsDocument8 pagesEffect of Intermittent Fasting On Brain Neurotransmitters, Neutrophils Phagocytic Activity, and Histopathological Finding in Some Organs in RatsBrian ColbornePas encore d'évaluation
- Resident Stem Cells and Regenerative TherapyD'EverandResident Stem Cells and Regenerative TherapyRegina Coeli dos Santos GoldenbergPas encore d'évaluation
- Final Synthesis Paper - Dhruvi Patel 2021-2022 1Document25 pagesFinal Synthesis Paper - Dhruvi Patel 2021-2022 1api-536778656Pas encore d'évaluation
- Adrenal Disorders: Physiology, Pathophysiology and TreatmentD'EverandAdrenal Disorders: Physiology, Pathophysiology and TreatmentAlice C. LevinePas encore d'évaluation
- Effects of Dietary Selenium On Histopathological Changes and T Cells of Spleen in Broilers Exposed To Aflatoxin B1Document10 pagesEffects of Dietary Selenium On Histopathological Changes and T Cells of Spleen in Broilers Exposed To Aflatoxin B1ANGIE CAMILA CASTILLO LOPEZPas encore d'évaluation
- BexigaDocument7 pagesBexigaana claraPas encore d'évaluation
- A Procedure For Creating A Frailty Index Based On Deficis in Mice - Parks - 2012Document11 pagesA Procedure For Creating A Frailty Index Based On Deficis in Mice - Parks - 2012gon71Pas encore d'évaluation
- Leaky Gut Solved: The Real Causes and Real Solutions for Healing Leaky Gut SyndromeD'EverandLeaky Gut Solved: The Real Causes and Real Solutions for Healing Leaky Gut SyndromePas encore d'évaluation
- Collagen Hydrolysate For OA and OsteoporosisDocument12 pagesCollagen Hydrolysate For OA and OsteoporosisSusan FPas encore d'évaluation
- AnorexiaDocument14 pagesAnorexiafhenrique77Pas encore d'évaluation
- Alimentos Hormonales, Experienciales y NeuronalesDocument7 pagesAlimentos Hormonales, Experienciales y NeuronalesJuanPas encore d'évaluation
- Pcttiidaa1841393 PDFDocument2 pagesPcttiidaa1841393 PDFRegina Marhadi SoniPas encore d'évaluation
- Attachment - 1 Drawings & Technical InformationDocument47 pagesAttachment - 1 Drawings & Technical InformationRegina Marhadi SoniPas encore d'évaluation
- Skripsi KikiDocument1 pageSkripsi KikiRegina Marhadi SoniPas encore d'évaluation
- The Coefficient of Linear ExpansionDocument4 pagesThe Coefficient of Linear ExpansionRegina Marhadi SoniPas encore d'évaluation
- Soni CVDocument16 pagesSoni CVRegina Marhadi SoniPas encore d'évaluation
- Adult Dosing & UsesDocument7 pagesAdult Dosing & UsesRegina Marhadi SoniPas encore d'évaluation
- Obesity: Nutrition Department Medical School University of Sumatera UtaraDocument31 pagesObesity: Nutrition Department Medical School University of Sumatera UtaraRegina Marhadi SoniPas encore d'évaluation
- Care of The Mother, Child and Adolescent (Well Client) : Essential Newborn Care From Evidence To PracticeDocument3 pagesCare of The Mother, Child and Adolescent (Well Client) : Essential Newborn Care From Evidence To PracticeAlec AnonPas encore d'évaluation
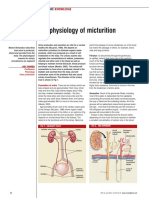
- 030722the Physiology of MicturitionDocument3 pages030722the Physiology of MicturitionAnnizah Paramitha100% (1)
- Test Ex.1 Read The Text. For Each Question, Choose The Best Answer (A, B, C or D) - (10 Points) Bird ResearcherDocument5 pagesTest Ex.1 Read The Text. For Each Question, Choose The Best Answer (A, B, C or D) - (10 Points) Bird ResearcherAndrei KazmarurkovPas encore d'évaluation
- Chitale Dairy Uses RFID To Improve Milk YieldsDocument5 pagesChitale Dairy Uses RFID To Improve Milk YieldsBhanu PathalaPas encore d'évaluation
- Forgotten Realms Archetypes Savagery & Shadow (11293249) PDFDocument55 pagesForgotten Realms Archetypes Savagery & Shadow (11293249) PDFMark Avrit100% (12)
- Elicityl - OligoTech - Human Milk Oligosaccharides HMOs - 2011v2Document4 pagesElicityl - OligoTech - Human Milk Oligosaccharides HMOs - 2011v2pascalsalvaPas encore d'évaluation
- Soal Big Kelas Xi SMT GenapDocument7 pagesSoal Big Kelas Xi SMT Genapariesta forestyaniPas encore d'évaluation
- A Review Ot Esthetic Pontic Design OptionsDocument12 pagesA Review Ot Esthetic Pontic Design OptionsAlejandro García Armenta100% (1)
- The 4x4 MatrixDocument23 pagesThe 4x4 Matrixpfi_jenPas encore d'évaluation
- DX Roleplaying HandbookDocument520 pagesDX Roleplaying HandbookJeffrey Lee89% (9)
- Answers Lab1 TerminologyDocument4 pagesAnswers Lab1 TerminologyColleen Mae MaciasPas encore d'évaluation
- Joyway Std-3-19Document40 pagesJoyway Std-3-19Vharry Watson33% (3)
- Ustrasana - Camel Pose - Yoga International PDFDocument8 pagesUstrasana - Camel Pose - Yoga International PDFToreØrnPas encore d'évaluation
- Flystrike Among Domestic Animals: An OverviewDocument3 pagesFlystrike Among Domestic Animals: An OverviewInternational Journal of Innovative Science and Research TechnologyPas encore d'évaluation
- Assesment C&G Prepare, Cook and Finish EggsDocument6 pagesAssesment C&G Prepare, Cook and Finish Eggsعلي صالحPas encore d'évaluation
- Rumen MicrofloraDocument14 pagesRumen MicrofloraDrRameem BlochPas encore d'évaluation
- GrammarDocument12 pagesGrammarMuhamad Zaini MasryPas encore d'évaluation
- English Paper 2 Form 3 Term 1 Exam 2017Document5 pagesEnglish Paper 2 Form 3 Term 1 Exam 2017Godfrey MuchaiPas encore d'évaluation
- Two - Four - Six - Eight - One - Eight: I. Choose The Correct Answer by Crossing (X) A, B or C!Document5 pagesTwo - Four - Six - Eight - One - Eight: I. Choose The Correct Answer by Crossing (X) A, B or C!Madi AlhajjPas encore d'évaluation
- Nervous System Lab ReportDocument3 pagesNervous System Lab Reportapi-296589906Pas encore d'évaluation
- Mordheim Treasure Hunt V2Document54 pagesMordheim Treasure Hunt V2Christian Camacho100% (1)
- English TestDocument13 pagesEnglish TestDJ MRPas encore d'évaluation
- RAW Lesson 04 - Patterns of DevelopmentDocument62 pagesRAW Lesson 04 - Patterns of DevelopmentArlance Sandra Marie MedinaPas encore d'évaluation
- Bobcat Classification and EvolutionDocument3 pagesBobcat Classification and EvolutionagnaPas encore d'évaluation
- Connect (4) New QuestionDocument18 pagesConnect (4) New QuestionGeorge ElsabaPas encore d'évaluation
- Animal Cytology (Compatibility Mode)Document13 pagesAnimal Cytology (Compatibility Mode)AdarshBijapurPas encore d'évaluation
- Animal Farm (Novel Units)Document8 pagesAnimal Farm (Novel Units)M M100% (1)
- Akhal-Teke (Horse)Document11 pagesAkhal-Teke (Horse)Amy KlezanPas encore d'évaluation
- Birds of Van ViharDocument8 pagesBirds of Van ViharDurgesh Kumar SinghPas encore d'évaluation
- ITC HS Export Policy 2018 PDFDocument392 pagesITC HS Export Policy 2018 PDFSachin AhujaPas encore d'évaluation