Vous aimerez peut-être aussi
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5795)
- Language Assessment in Year 2 KSSRDocument13 pagesLanguage Assessment in Year 2 KSSRCahaya Hati100% (1)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- CH 1 MCQ ETIDocument14 pagesCH 1 MCQ ETIShlok PurohitPas encore d'évaluation
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- 731 Mitendra Pratap SinghDocument2 pages731 Mitendra Pratap Singhmitendra pratap singhPas encore d'évaluation
- Michael Buble - HomeDocument5 pagesMichael Buble - Homepurbaalam0702Pas encore d'évaluation
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- Geotecnical Limit State AustralianDocument6 pagesGeotecnical Limit State Australianprashanth c.nPas encore d'évaluation
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- OFBiz POS User Manual - Version 9.11Document24 pagesOFBiz POS User Manual - Version 9.11shibubabu123Pas encore d'évaluation
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- ISO 261 EN 2010 - Page2 PDFDocument1 pageISO 261 EN 2010 - Page2 PDFd3bilPas encore d'évaluation
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- F 8 T 161 P 960 N 1Document7 pagesF 8 T 161 P 960 N 1Jessica PratesPas encore d'évaluation
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- English For Military PersonnelDocument9 pagesEnglish For Military PersonnelGilang Satia LaksanaPas encore d'évaluation
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- PD083 05Document1 pagePD083 05Christian Linares AbreuPas encore d'évaluation
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (400)
- PTE Academic - Strategies For Summarize The Written TextDocument9 pagesPTE Academic - Strategies For Summarize The Written TextPradeep PaudelPas encore d'évaluation
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- Eatclub D5Z1X6Document1 pageEatclub D5Z1X6Devansh nayakPas encore d'évaluation
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (74)
- Ayaz Ahmed SoomroDocument9 pagesAyaz Ahmed SoomroNoman RathorePas encore d'évaluation
- RoyalBritishBank V TurquanDocument3 pagesRoyalBritishBank V TurquanAlbukhari AliasPas encore d'évaluation
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- MIL-H-6875H Heat Treatment of SteelDocument29 pagesMIL-H-6875H Heat Treatment of Steelgoathead61Pas encore d'évaluation
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (345)
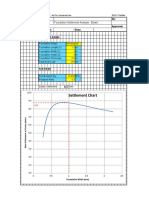
- Settlement Chart: Foundation Settlement Analysis - ElasticDocument2 pagesSettlement Chart: Foundation Settlement Analysis - ElasticCSEC Uganda Ltd.Pas encore d'évaluation
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- IoT-Based Water Monitoring SystemDocument5 pagesIoT-Based Water Monitoring SystemManuel Luis Delos SantosPas encore d'évaluation
- Sharp+Lc c6554u+Difference+Lc 65d64uDocument30 pagesSharp+Lc c6554u+Difference+Lc 65d64uNicholas RileyPas encore d'évaluation
- WASA Wastewater and Potable Water Design Requirements PDFDocument233 pagesWASA Wastewater and Potable Water Design Requirements PDFYassin AlkadyPas encore d'évaluation
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- 2018 2019UndergraduateEnglishTaughtCoursesDocument40 pages2018 2019UndergraduateEnglishTaughtCoursesmalu accountPas encore d'évaluation
- Comparative and Superlative Adjectives Fun Activities Games Grammar Guides 10529Document14 pagesComparative and Superlative Adjectives Fun Activities Games Grammar Guides 10529Nhung Mai ThịPas encore d'évaluation
- Anh 7 Cum 1 (2017-2018)Document6 pagesAnh 7 Cum 1 (2017-2018)anhthunoban2011Pas encore d'évaluation
- MBA Project Report of DY Patil Distance MBADocument4 pagesMBA Project Report of DY Patil Distance MBAPrakashB144Pas encore d'évaluation
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1091)
- Questions PetrobowlDocument58 pagesQuestions PetrobowlCuloPas encore d'évaluation
- Resume ManikandanDocument3 pagesResume ManikandanPMANIKANDANSVNPas encore d'évaluation
- Federico Giusfredi - Syntax of The Luwian LanguageDocument229 pagesFederico Giusfredi - Syntax of The Luwian LanguageLucySky7Pas encore d'évaluation
- Connector GuideDocument20 pagesConnector GuideBastian VarelaPas encore d'évaluation
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- Perfect Email FormulaDocument4 pagesPerfect Email FormulaBallpoitAryanPas encore d'évaluation
- Marketing Plan Rental LaptopDocument15 pagesMarketing Plan Rental LaptopDany AkbarPas encore d'évaluation
- GL2012 - 001-High Volume Short Stay Surgery Model ToolkitDocument30 pagesGL2012 - 001-High Volume Short Stay Surgery Model ToolkitPankaj MittalPas encore d'évaluation
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)