Vous aimerez peut-être aussi
- Marco TeóricoDocument3 pagesMarco TeóricoAna Cristina Ayllón Velarde100% (1)
- Biopotenciales: Impulso nervioso y potencialesDocument29 pagesBiopotenciales: Impulso nervioso y potencialesYakeline VelasquezPas encore d'évaluation
- Conducción Del Impulso NerviosoDocument17 pagesConducción Del Impulso NerviosoAlbertina Klüver GrossPas encore d'évaluation
- El Origen de Los BiopotencialesDocument8 pagesEl Origen de Los BiopotencialesLY Ariel HaroldPas encore d'évaluation
- Conduccion Del Impulso NerviosoDocument17 pagesConduccion Del Impulso NerviosoomarcvetPas encore d'évaluation
- Impulso Nervioso y SinapsisDocument10 pagesImpulso Nervioso y SinapsisJesus García QuezadaPas encore d'évaluation
- BiopotencialesDocument241 pagesBiopotencialesHéctor Alan AguilarPas encore d'évaluation
- 1 Seminario de Fisiologia ResueltoDocument30 pages1 Seminario de Fisiologia ResueltoJorgeMaquiNarcisoPas encore d'évaluation
- UTI Práctica5. Propiedades Pasivas y Activas de Las Células Excitables 2Document12 pagesUTI Práctica5. Propiedades Pasivas y Activas de Las Células Excitables 2Luis Enrique MarquezPas encore d'évaluation
- MONOGRAFIA DE PotencialesDocument24 pagesMONOGRAFIA DE PotencialesKilleer AguilarPas encore d'évaluation
- Taller Potencial de AcciónDocument3 pagesTaller Potencial de AcciónDaniellaPas encore d'évaluation
- Neuronal Excitación y ConducciónDocument13 pagesNeuronal Excitación y ConducciónCésar HaroPas encore d'évaluation
- Investigaciones para Fisio en Orden CronologicoDocument12 pagesInvestigaciones para Fisio en Orden CronologicoMiguel AmbrocioPas encore d'évaluation
- Historia y efectos de la electroterapia médicaDocument14 pagesHistoria y efectos de la electroterapia médicaMarcelo Vera SuasnávarPas encore d'évaluation
- Voltaje Clamp: Descripción y aplicaciones en instrumentación biomédicaDocument3 pagesVoltaje Clamp: Descripción y aplicaciones en instrumentación biomédicaJonathan YamberlaPas encore d'évaluation
- Seminario Fisiología - BiopotencialesDocument16 pagesSeminario Fisiología - BiopotencialesAdriana Vivar BalcazarPas encore d'évaluation
- Informe 1 - FisiologíaDocument11 pagesInforme 1 - FisiologíaAngela SanchezPas encore d'évaluation
- Simulación PA modelo HHDocument8 pagesSimulación PA modelo HHbryanPas encore d'évaluation
- Guia Potencial de Accion y NeurotransmisoresDocument11 pagesGuia Potencial de Accion y NeurotransmisoresAlvaro Andres Sanchez HurtadoPas encore d'évaluation
- TP 4 PropiedadespasivasdelamembranaDocument10 pagesTP 4 PropiedadespasivasdelamembranaVero OrtegaPas encore d'évaluation
- BiopotencialesDocument2 pagesBiopotencialesadrimedi15Pas encore d'évaluation
- Seminario de PotencialesDocument14 pagesSeminario de PotencialesAlexanther Palomino ReateguiPas encore d'évaluation
- Electrodos PDFDocument43 pagesElectrodos PDFTulio Andrés Oviedo VargasPas encore d'évaluation
- BiopotencialesDocument6 pagesBiopotencialesLiseth100% (1)
- Teoría Del Potencial de Acción de La Membrana Postulado Por Lloyd Hodgkin y Andrew Fielding HuxleyDocument7 pagesTeoría Del Potencial de Acción de La Membrana Postulado Por Lloyd Hodgkin y Andrew Fielding HuxleyBrenPas encore d'évaluation
- ELECTROMEDICINADocument20 pagesELECTROMEDICINAKarol FollecoPas encore d'évaluation
- INFORMEDocument14 pagesINFORMESara Pancorbo VargasPas encore d'évaluation
- Potenciales membrana-40Document19 pagesPotenciales membrana-40Andy Acevedo50% (2)
- Seminario de Fisiología PotencialesDocument21 pagesSeminario de Fisiología PotencialesEdwin CallePas encore d'évaluation
- Biofisica de Las Membranas ExcitablesDocument19 pagesBiofisica de Las Membranas ExcitablesYamil Valencia100% (1)
- Seminario de Fisiologia 01Document16 pagesSeminario de Fisiologia 01Stephanie SanchezPas encore d'évaluation
- GTP 2 Un 1 2MDocument19 pagesGTP 2 Un 1 2Mli_sisayPas encore d'évaluation
- Electrofisiología cardíacaDocument12 pagesElectrofisiología cardíacaAlejandro Pari VillenaPas encore d'évaluation
- Tarea 2 - Trabajo ColaborativoDocument46 pagesTarea 2 - Trabajo ColaborativoJuan Carlos Grisales MenesesPas encore d'évaluation
- Ee - Grupo 2Document5 pagesEe - Grupo 2Dayana Alexandra Palomino AlbinesPas encore d'évaluation
- BioseñalesDocument29 pagesBioseñalesmanuel.corrales100% (1)
- Capitulo 3 - Neurociencia PDFDocument16 pagesCapitulo 3 - Neurociencia PDFDavid RojasPas encore d'évaluation
- Seminario de Potenciales FisiologiaDocument10 pagesSeminario de Potenciales FisiologiaGuillermo Ruiz Cabrera0% (1)
- 3 Bases ElectricasDocument8 pages3 Bases ElectricasMaría José FernándezPas encore d'évaluation
- Potenciales Bioeléctricos USEP 2022Document28 pagesPotenciales Bioeléctricos USEP 2022YESENIA SARAIT AGUILAR HERRERAPas encore d'évaluation
- Tejidos excitables y potencial de acciónDocument38 pagesTejidos excitables y potencial de acciónFrancisco CermeñoPas encore d'évaluation
- BioelectricidadDocument8 pagesBioelectricidadJesus AstorPas encore d'évaluation
- Diapositivas Conduccion Del Impulso y Excitabilidad CelularDocument60 pagesDiapositivas Conduccion Del Impulso y Excitabilidad Celularkirys18Pas encore d'évaluation
- 1er SEMINARIO BIOPOTENCIALESDocument15 pages1er SEMINARIO BIOPOTENCIALESHannah Barlow100% (1)
- Vídeos Explicación Unidad Iii (Fisiología)Document6 pagesVídeos Explicación Unidad Iii (Fisiología)Amy Lilian Verdooren UzcateguiPas encore d'évaluation
- Emg & EegDocument10 pagesEmg & EegManuel AlonsoPas encore d'évaluation
- Fisi 5Document5 pagesFisi 5AlexPas encore d'évaluation
- CA 6 Fisica Medica - En.espanolDocument50 pagesCA 6 Fisica Medica - En.espanolCristhian Alexander Rafael BaylònPas encore d'évaluation
- BIOELECTRICIDAD FisicaDocument6 pagesBIOELECTRICIDAD FisicaAnngie Lisbeth Requejo MasPas encore d'évaluation
- Seminario Biopotenciales FisiologiaDocument9 pagesSeminario Biopotenciales Fisiologiayamirjossue100% (9)
- BiopotencialesDocument20 pagesBiopotencialesRoland CaixaPas encore d'évaluation
- Practica 6. Potencial de Acción CompuestoDocument12 pagesPractica 6. Potencial de Acción CompuestoErnesto BecerroPas encore d'évaluation
- Psicofisiologia 2Document11 pagesPsicofisiologia 2Juan Estarling TrinidadPas encore d'évaluation
- Resumen de El Principio de Incertidumbre de Stephen Hawking: RESÚMENES UNIVERSITARIOSD'EverandResumen de El Principio de Incertidumbre de Stephen Hawking: RESÚMENES UNIVERSITARIOSÉvaluation : 5 sur 5 étoiles5/5 (1)
- El problema del aumento de la energía humana (Traducido): Con referencias especiales al aprovechamiento de la energía solarD'EverandEl problema del aumento de la energía humana (Traducido): Con referencias especiales al aprovechamiento de la energía solarPas encore d'évaluation
- Campos Electromagnéticos: La exposición no es optativa, nadie puede escapar a ellaD'EverandCampos Electromagnéticos: La exposición no es optativa, nadie puede escapar a ellaPas encore d'évaluation
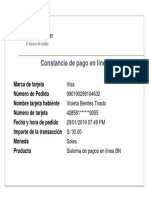
- Visa 990190299104632 PDFDocument1 pageVisa 990190299104632 PDFSeleny Soto BenitesPas encore d'évaluation
- Adaptación, Validación y Estandarización de Cuestionarios para Egresados y Empleadores en El Estado de MéxicoDocument19 pagesAdaptación, Validación y Estandarización de Cuestionarios para Egresados y Empleadores en El Estado de MéxicoKatherine PradoPas encore d'évaluation
- Comunicacion AsertivaDocument33 pagesComunicacion AsertivaMartin Vidal RealEstatePas encore d'évaluation
- Anamnesis PsicologicaDocument20 pagesAnamnesis PsicologicaAdrian Castro100% (1)
- Beck DepresionDocument12 pagesBeck DepresionLorena Soledad BazánPas encore d'évaluation
- Dialnet MujeresMaltratadasEnLaActualidadApuntesDesdeLaClin 5164625Document8 pagesDialnet MujeresMaltratadasEnLaActualidadApuntesDesdeLaClin 5164625licnoeliaPas encore d'évaluation
- Los Celos Robert Briceno PDFDocument12 pagesLos Celos Robert Briceno PDFSeleny Soto BenitesPas encore d'évaluation
- Resiliencia y Dolor CrónicoDocument11 pagesResiliencia y Dolor CrónicoSeleny Soto BenitesPas encore d'évaluation
- Técnica de RetribuciónDocument5 pagesTécnica de RetribuciónSeleny Soto BenitesPas encore d'évaluation
- Mindfulness, ansiedad y estudiantes universitariosDocument43 pagesMindfulness, ansiedad y estudiantes universitariosSeleny Soto BenitesPas encore d'évaluation
- Escala Autonoma SADocument6 pagesEscala Autonoma SAMaribel DelaordencazorlaPas encore d'évaluation
- 210 758 1 PBDocument22 pages210 758 1 PBcaudilloazulPas encore d'évaluation
- E-MAIL LABORAL: Facultades y Limitaciones Del EmpleadorDocument8 pagesE-MAIL LABORAL: Facultades y Limitaciones Del EmpleadorFidel Francisco Quispe HinostrozaPas encore d'évaluation
- Trastorno Depresivo Mayor y AgorafobiaDocument111 pagesTrastorno Depresivo Mayor y AgorafobiaSeleny Soto BenitesPas encore d'évaluation
- Guía para Padres y MadresDocument72 pagesGuía para Padres y MadresSeleny Soto BenitesPas encore d'évaluation
- Gestión Del Talento HumanoDocument88 pagesGestión Del Talento HumanoSeleny Soto Benites100% (1)
- Formato de TernaDocument3 pagesFormato de TernaSeleny Soto Benites100% (2)
- Guia Pacientes Geriatricos PDFDocument209 pagesGuia Pacientes Geriatricos PDFauroraPas encore d'évaluation
- Ines Temple. Usted S.A.Document70 pagesInes Temple. Usted S.A.Luis Barrera100% (4)
- Salud Sexual y Reproductiva Adolescente y JuvenilDocument429 pagesSalud Sexual y Reproductiva Adolescente y JuvenilSeleny Soto BenitesPas encore d'évaluation
- Kinsey para HombresDocument2 pagesKinsey para HombresSeleny Soto BenitesPas encore d'évaluation
- Módulo de Psicología EducativaDocument159 pagesMódulo de Psicología EducativaSeleny Soto BenitesPas encore d'évaluation
- PARAFILIASDocument58 pagesPARAFILIASSeleny Soto Benites100% (3)
- Sexismo: Infravaloración o InfrahumanizaciónDocument21 pagesSexismo: Infravaloración o InfrahumanizaciónSeleny Soto BenitesPas encore d'évaluation
- Kinsey MujeresDocument2 pagesKinsey MujeresSeleny Soto BenitesPas encore d'évaluation
- Manual de Psicopatologia2013Document10 pagesManual de Psicopatologia2013polariz3033% (3)
- Liderazgo EnsayoDocument62 pagesLiderazgo EnsayoSeleny Soto Benites100% (1)
- Escala de Dependencia EmocionalDocument2 pagesEscala de Dependencia EmocionalSeleny Soto BenitesPas encore d'évaluation
- Protocolo Bas 3Document8 pagesProtocolo Bas 3Seleny Soto Benites100% (1)
- Manual Del Bas-3Document12 pagesManual Del Bas-3Seleny Soto Benites86% (35)
- Carbohidratos y lípidosDocument11 pagesCarbohidratos y lípidosBRIGITTE URQUIJOPas encore d'évaluation
- Guia 9 Impulso Momentum Conservacion Momentum de Un Sistema Aislado s1 2013 2Document3 pagesGuia 9 Impulso Momentum Conservacion Momentum de Un Sistema Aislado s1 2013 2Bárbara Muñoz Madariaga0% (1)
- Informe 1 CSDocument10 pagesInforme 1 CSSara Rojas AgudeloPas encore d'évaluation
- Energía necesaria para separar iones en una red cristalinaDocument10 pagesEnergía necesaria para separar iones en una red cristalinaKibor Coronel CruzPas encore d'évaluation
- Mezcl y AleDocument4 pagesMezcl y AleFrank Alex RamirezPas encore d'évaluation
- Informe 2da Ley de NewtonDocument13 pagesInforme 2da Ley de NewtonJonathan BatistaPas encore d'évaluation
- Hoja de Seguridad Aditivo SikaDocument7 pagesHoja de Seguridad Aditivo SikaEmiro OrozcoPas encore d'évaluation
- Vitrina Manual de Usuario PDFDocument1 pageVitrina Manual de Usuario PDFDamianPas encore d'évaluation
- Cristalización: proceso, teorías y equiposDocument28 pagesCristalización: proceso, teorías y equiposDavid Hawmann Diaz50% (2)
- Radiactividad PDFDocument22 pagesRadiactividad PDFcereplPas encore d'évaluation
- 2022-3 Estructura CristalinaxDocument28 pages2022-3 Estructura CristalinaxGonza SavorPas encore d'évaluation
- 1 QUÍMICA (1-6) .Indd cs5Document66 pages1 QUÍMICA (1-6) .Indd cs5EfrainTamaraPas encore d'évaluation
- Manifestaciones de La EnergíaDocument5 pagesManifestaciones de La EnergíaAbril SanchezPas encore d'évaluation
- TESIS Jiménez CotillasDocument416 pagesTESIS Jiménez CotillasNeyda Flores VasquezPas encore d'évaluation
- Genreador de HidrogenoDocument16 pagesGenreador de HidrogenoLeonardoDavidMezuLobatoPas encore d'évaluation
- Acidos, Bases E Indicadores: Reporte: Practica de Laboratorio 2Document20 pagesAcidos, Bases E Indicadores: Reporte: Practica de Laboratorio 2Gisell MartinezPas encore d'évaluation
- Actividad SALESDocument3 pagesActividad SALESSandra Yecenia AragonPas encore d'évaluation
- Paper Practica 4 ElectroDocument10 pagesPaper Practica 4 Electrokate vilcapaza Charca100% (1)
- Resumen Equilibrio IónicoDocument6 pagesResumen Equilibrio IónicoHéctor Portilla50% (2)
- DisolucionesDocument4 pagesDisolucionesAlberto ManzanoPas encore d'évaluation
- Análisis de Deformaciones Elásticas de Estructuras.Document12 pagesAnálisis de Deformaciones Elásticas de Estructuras.Manuel ColmenaresPas encore d'évaluation
- EnzimasDocument44 pagesEnzimasPedro Chirre AlfaroPas encore d'évaluation
- Trabajo PR Actico N 3 Aireh UmedoDocument2 pagesTrabajo PR Actico N 3 Aireh UmedoLENDPas encore d'évaluation
- Ejercicios Análisis InstrumentalDocument4 pagesEjercicios Análisis InstrumentalMarcos JimenezPas encore d'évaluation
- FTC-580 Rev. ADocument3 pagesFTC-580 Rev. Ajlop57e8r78Pas encore d'évaluation
- Sìlabo Del Curso de Quìmica de Los PolìmerosDocument7 pagesSìlabo Del Curso de Quìmica de Los PolìmerosRosmeri DelgadoPas encore d'évaluation
- 33 34 Electrotecnia 1Document12 pages33 34 Electrotecnia 1jostin berronesPas encore d'évaluation
- Espectroscopia UV-Vis: Transiciones electrónicas y aplicacionesDocument27 pagesEspectroscopia UV-Vis: Transiciones electrónicas y aplicacionesGABRIEL MARINO MARTINEZ PARRAPas encore d'évaluation
- Guia Enlace QuimicoDocument8 pagesGuia Enlace QuimicoFoXD DebugPas encore d'évaluation
- Dilatación térmica: conceptos básicosDocument55 pagesDilatación térmica: conceptos básicosdiego alejandro leon zegarraPas encore d'évaluation