Vous aimerez peut-être aussi
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (400)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (74)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (345)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Auditory PathwayDocument16 pagesAuditory PathwayDrsreeram ValluriPas encore d'évaluation
- Auditory Pathways: Anatomy and Physiology: Handbook of Clinical Neurology March 2015Document24 pagesAuditory Pathways: Anatomy and Physiology: Handbook of Clinical Neurology March 2015florensiaPas encore d'évaluation
- Clin Aud I HandoutDocument48 pagesClin Aud I HandoutelFadhlyPas encore d'évaluation
- 24 Inner Ear Anatomy, Emberyology, Drugs Ototoxicity (Done)Document24 pages24 Inner Ear Anatomy, Emberyology, Drugs Ototoxicity (Done)osamaePas encore d'évaluation
- Central Auditory PathwaysDocument31 pagesCentral Auditory Pathwayslion2chPas encore d'évaluation
- Reflex Measurements: Quick GuideDocument10 pagesReflex Measurements: Quick GuidefilippobPas encore d'évaluation
- James O. Pickles - An Introduction To The Physiology of Hearing-Emerald Group Publishing Limited (2012)Document459 pagesJames O. Pickles - An Introduction To The Physiology of Hearing-Emerald Group Publishing Limited (2012)mane100% (1)
- 1 s2.0 S0306452214009932 MainDocument12 pages1 s2.0 S0306452214009932 MainRubén Alcalá ArgüellesPas encore d'évaluation
- Cranial Nerve 8 (Vestibulo-Cochlear)Document14 pagesCranial Nerve 8 (Vestibulo-Cochlear)osakaPas encore d'évaluation
- Physio HearingDocument56 pagesPhysio Hearing65 Nidheesh Kumar PrabakamoorthyPas encore d'évaluation
- Acoustic Reflex Threshold (ART) Patterns: An Interpretation Guide For Students and Supervisors: Course Material and Exam Questions Course MaterialDocument17 pagesAcoustic Reflex Threshold (ART) Patterns: An Interpretation Guide For Students and Supervisors: Course Material and Exam Questions Course Materialn_almagroPas encore d'évaluation
- Auditory PathwayDocument33 pagesAuditory PathwayAnusha Varanasi100% (1)
- Foundations of Behavioral Neuroscience 9th Edition Carlson Test BankDocument29 pagesFoundations of Behavioral Neuroscience 9th Edition Carlson Test Bankxeniaeira98897% (34)
- Masking Level Difference and ElectrophysiologicalDocument8 pagesMasking Level Difference and ElectrophysiologicalFernanda CardozoPas encore d'évaluation
- Physiology of The Auditory SystemDocument47 pagesPhysiology of The Auditory Systemmkct111Pas encore d'évaluation
- Every Brain Needs Music The Neuroscience of Making and Listening To MusicDocument289 pagesEvery Brain Needs Music The Neuroscience of Making and Listening To MusicJorge Antonio Pichardo100% (7)
- Auditory ReceptorsDocument15 pagesAuditory ReceptorsJeanette RiosPas encore d'évaluation
- Health Aspects of Wireless Communication: Auditory Perception of Microwaves - Hearing Microwaves by James C. LinDocument4 pagesHealth Aspects of Wireless Communication: Auditory Perception of Microwaves - Hearing Microwaves by James C. LinbingwazzupPas encore d'évaluation
- NEURO Ch. 1-5 Quiz 1 NotesDocument31 pagesNEURO Ch. 1-5 Quiz 1 NotesMelissa OrtizPas encore d'évaluation
- UntitledDocument378 pagesUntitledLaurent BarbierPas encore d'évaluation
- Cap 11 - 22 Handbook of Clinical AudiologyDocument216 pagesCap 11 - 22 Handbook of Clinical AudiologyCami IrribarraPas encore d'évaluation
- Hearing Physiology - GuytonDocument11 pagesHearing Physiology - GuytonDuc HaPas encore d'évaluation
- Auditory PerceptionDocument29 pagesAuditory PerceptionRose RodziPas encore d'évaluation
- The First Central Auditory Structure: The Cochlear NucleusDocument56 pagesThe First Central Auditory Structure: The Cochlear NucleusAlison LuizPas encore d'évaluation
- (Progress in Brain Research 166) B. Langguth, G. Hajak, T. Kleinjung, A. Cacace and A.R. Møller (Eds.) - Tinnitus - Pathophysiology and Treatment (2007, Elsevier Science)Document511 pages(Progress in Brain Research 166) B. Langguth, G. Hajak, T. Kleinjung, A. Cacace and A.R. Møller (Eds.) - Tinnitus - Pathophysiology and Treatment (2007, Elsevier Science)fahuroPas encore d'évaluation
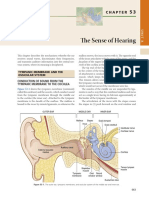
- The Sense of HearingDocument11 pagesThe Sense of HearingAfdalia AmandaPas encore d'évaluation
- The Dangers of Edosing With Binaural Beats: PathwaysDocument2 pagesThe Dangers of Edosing With Binaural Beats: PathwaysAgustina IturriPas encore d'évaluation
- 12b. The Central Auditory PathwayDocument2 pages12b. The Central Auditory PathwayShivan UmamaheswaranPas encore d'évaluation
- Sound Localization EssayDocument11 pagesSound Localization EssayscholifyPas encore d'évaluation
- Acoustic - Stapedius - Reflexes PDFDocument6 pagesAcoustic - Stapedius - Reflexes PDFSOFTMERCURYPas encore d'évaluation