Vous aimerez peut-être aussi
- The Handbook of Histopathological Practices in Aquatic Environments: Guide to Histology for Environmental ToxicologyD'EverandThe Handbook of Histopathological Practices in Aquatic Environments: Guide to Histology for Environmental ToxicologyÉvaluation : 4.5 sur 5 étoiles4.5/5 (2)
- J. Bacteriol.-1980-Ingram-481-8Document9 pagesJ. Bacteriol.-1980-Ingram-481-8Phytomenadione La Vitamine KPas encore d'évaluation
- Universidad Nacional de Colombia, A.A. 14490, Bogot A D. C., ColombiaDocument1 pageUniversidad Nacional de Colombia, A.A. 14490, Bogot A D. C., ColombiaDanielRicardoDelgadoPas encore d'évaluation
- Journal of Agricultural and Food Chemistry Volume 49 Issue 6 2001 (Doi 10.1021 - jf001413m) Burda, Stanislaw Oleszek, Wieslaw - Antioxidant and Antiradical Activities of FlavonoidsDocument6 pagesJournal of Agricultural and Food Chemistry Volume 49 Issue 6 2001 (Doi 10.1021 - jf001413m) Burda, Stanislaw Oleszek, Wieslaw - Antioxidant and Antiradical Activities of FlavonoidsnaelaniesaPas encore d'évaluation
- Antibacterial Action 2-Bromo-2-Nitropropane-1,3-Diol (Bronopol)Document6 pagesAntibacterial Action 2-Bromo-2-Nitropropane-1,3-Diol (Bronopol)lokesh1331Pas encore d'évaluation
- Fisher Et Al-2009-Journal of Applied MicrobiologyDocument7 pagesFisher Et Al-2009-Journal of Applied MicrobiologydigdouwPas encore d'évaluation
- Investigation of Acetone-Butanol-Ethanol (ABE) Fermentation by FluorescenceDocument4 pagesInvestigation of Acetone-Butanol-Ethanol (ABE) Fermentation by FluorescencechelogkPas encore d'évaluation
- Cacciatore in VivoDocument11 pagesCacciatore in VivoClauTorePas encore d'évaluation
- Bacterial Degradation Compounds: of DetergentDocument6 pagesBacterial Degradation Compounds: of Detergentp_srib4uPas encore d'évaluation
- Biological Activity and Variation of Toxicity of Probenecid-IJACS-2018Document7 pagesBiological Activity and Variation of Toxicity of Probenecid-IJACS-2018Soumen BhattacharjeePas encore d'évaluation
- Athyrium Multidentatum: Antioxidant Activity of Polysaccharides Extracted From (Doll) ChingDocument6 pagesAthyrium Multidentatum: Antioxidant Activity of Polysaccharides Extracted From (Doll) ChingmeriemPas encore d'évaluation
- Effect of Different Non-Ionic Surfactants On The BDocument10 pagesEffect of Different Non-Ionic Surfactants On The BFatma WatiPas encore d'évaluation
- Indicator OrganismsDocument10 pagesIndicator Organismsrocky21stPas encore d'évaluation
- Human Pharmaceuticals in Wastewater TreaDocument27 pagesHuman Pharmaceuticals in Wastewater TreaM Irfan KhanPas encore d'évaluation
- Cyanobacteria LDocument7 pagesCyanobacteria LdhiraniaPas encore d'évaluation
- Vitamina CDocument4 pagesVitamina CChoco LegUePas encore d'évaluation
- 1950 - Edsall Et Al. - Light Scattering in Solutions of Serum Albumin Effects of Charge and IonicDocument16 pages1950 - Edsall Et Al. - Light Scattering in Solutions of Serum Albumin Effects of Charge and IonicNayara CarvalhoPas encore d'évaluation
- Principles of Toxicology: The Study of PoisonsDocument36 pagesPrinciples of Toxicology: The Study of PoisonsKedrine AcengPas encore d'évaluation
- Chatterjee 2006Document10 pagesChatterjee 2006Yunita MarwahPas encore d'évaluation
- Dez Oxo Rub IcinDocument4 pagesDez Oxo Rub Icinalex_andra_22Pas encore d'évaluation
- Rodriguez Pez Cebra 3BP-2015Document5 pagesRodriguez Pez Cebra 3BP-2015Gabriela RodriguezPas encore d'évaluation
- Effects of Solvents and Surfactant Agents On The Female and Larvae of Cattle Tick Boophilus MicroplusDocument4 pagesEffects of Solvents and Surfactant Agents On The Female and Larvae of Cattle Tick Boophilus MicroplusIsabela SantosPas encore d'évaluation
- Baun 2008Document9 pagesBaun 2008Luis RamosPas encore d'évaluation
- Zaks and Klibanov, 1984 PDFDocument3 pagesZaks and Klibanov, 1984 PDFalvaro_cruzPas encore d'évaluation
- The Allelopathy and Allelopathic Mechanism of Phenolic Acids On Toxic Microcystis AeruginosaDocument7 pagesThe Allelopathy and Allelopathic Mechanism of Phenolic Acids On Toxic Microcystis AeruginosaArmando Chiclla SalazarPas encore d'évaluation
- LC MS EstrogenDocument11 pagesLC MS EstrogenHà NgọcPas encore d'évaluation
- Estudio de Ecotoxicidad y Genotoxicidad en Recursos Hídricos - IisDocument8 pagesEstudio de Ecotoxicidad y Genotoxicidad en Recursos Hídricos - IisĹIZBETH VARGAS SANTIVAÑEZPas encore d'évaluation
- 2016 - Guo Et Al. Effects of Antibiotics On The Growth and Physiology of Chlorophytes, Cyanobacteria and A Diatom PDFDocument14 pages2016 - Guo Et Al. Effects of Antibiotics On The Growth and Physiology of Chlorophytes, Cyanobacteria and A Diatom PDFSacra PsyntergiaPas encore d'évaluation
- Development, Validation, and Applications of A New Laboratory-Scale Indirect Impedancemeter For Rapid Microbial ControlDocument7 pagesDevelopment, Validation, and Applications of A New Laboratory-Scale Indirect Impedancemeter For Rapid Microbial ControloldpenguinPas encore d'évaluation
- Benzene BEIDocument14 pagesBenzene BEIsoehartonoPas encore d'évaluation
- 18 PDFDocument6 pages18 PDFТатьяна СаенкоPas encore d'évaluation
- Are Natural Deep Eutectic Solvents The Missing Link in Understanding Cellular Metabolism and Physiology?Document5 pagesAre Natural Deep Eutectic Solvents The Missing Link in Understanding Cellular Metabolism and Physiology?Emmanuel Avalos HuartePas encore d'évaluation
- LarissaDocument6 pagesLarissalharhasouzaPas encore d'évaluation
- Alicino 1946Document2 pagesAlicino 1946Gustavo D.B.Pas encore d'évaluation
- Colchicine Glycorandomization Influences Cytotoxicity and Mechanism of ActionDocument2 pagesColchicine Glycorandomization Influences Cytotoxicity and Mechanism of ActionLeonardo Antonio Castillo ZegarraPas encore d'évaluation
- Drilling Fluid Regulations Outside Latin AmericaDocument23 pagesDrilling Fluid Regulations Outside Latin AmericaMac HargitayPas encore d'évaluation
- Fems Microbiology Letters - 2006 - Schoefer - A Fluorescence Quenching Test For The Detection of Flavonoid TransformationDocument4 pagesFems Microbiology Letters - 2006 - Schoefer - A Fluorescence Quenching Test For The Detection of Flavonoid TransformationRodos LaboratoriesPas encore d'évaluation
- DEHP Induced Oxidative Stress Suppress Sperm Quality and Serum Androgen Concentration in Wistar RatsDocument8 pagesDEHP Induced Oxidative Stress Suppress Sperm Quality and Serum Androgen Concentration in Wistar RatsInternational Journal of Innovative Science and Research TechnologyPas encore d'évaluation
- Metabolic Engineering: Kyle A. Zingaro, Eleftherios Terry PapoutsakisDocument10 pagesMetabolic Engineering: Kyle A. Zingaro, Eleftherios Terry PapoutsakisKay WhitePas encore d'évaluation
- Chlorotoluene HPDocument71 pagesChlorotoluene HPShailesh LahotiPas encore d'évaluation
- Dioxido de CloroDocument12 pagesDioxido de CloroEli De La CruzPas encore d'évaluation
- Paper Claudio CB MAZDocument9 pagesPaper Claudio CB MAZClauTorePas encore d'évaluation
- Science of The Total Environment: Vera Homem, Arminda Alves, Lúcia SantosDocument9 pagesScience of The Total Environment: Vera Homem, Arminda Alves, Lúcia SantosLe Thuy Uyen NhaPas encore d'évaluation
- Chemistry Individual InvestigationDocument14 pagesChemistry Individual InvestigationAdrien G. S. WaldPas encore d'évaluation
- BAM: Enumeration of Escherichia Coli and The Coliform BacteriaDocument8 pagesBAM: Enumeration of Escherichia Coli and The Coliform BacteriatintfenPas encore d'évaluation
- 2008 Coats IdentificationTylosinDocument6 pages2008 Coats IdentificationTylosinbejlvwirsharklasersomPas encore d'évaluation
- Jurnal 10Document17 pagesJurnal 10Daniall AbdanPas encore d'évaluation
- Enumeration of Escherichia Coli and The Coliform BacteriaDocument11 pagesEnumeration of Escherichia Coli and The Coliform Bacteriaarku64Pas encore d'évaluation
- Effects of Leukotrienes On Susceptibility of The Rat Stomach To Damage and Investigation of The Mechanism of ActionDocument9 pagesEffects of Leukotrienes On Susceptibility of The Rat Stomach To Damage and Investigation of The Mechanism of ActionGeorgi GugicevPas encore d'évaluation
- Yasuda 1982Document6 pagesYasuda 1982Saarah KhairunnisaPas encore d'évaluation
- Schirmer 1999 UseDocument12 pagesSchirmer 1999 UseJoana RosaPas encore d'évaluation
- 2011 VERSÃO PUBLICADA Towards A Better Understanding of Ipomoea Asarifolia Toxicity - Evidence of The Involvement of A Leaf LectinDocument7 pages2011 VERSÃO PUBLICADA Towards A Better Understanding of Ipomoea Asarifolia Toxicity - Evidence of The Involvement of A Leaf Lectinnilberto2Pas encore d'évaluation
- Prolonged Bactericidal Effect of Sodium HypochloriteDocument3 pagesProlonged Bactericidal Effect of Sodium HypochloritegiuseppegnrPas encore d'évaluation
- 8.interaction of Methyl and Propyl Parabens With Selected Sucrose EstersDocument4 pages8.interaction of Methyl and Propyl Parabens With Selected Sucrose EstersÁnh NguyễnPas encore d'évaluation
- Lourenco 2012Document7 pagesLourenco 2012ichwan bagusPas encore d'évaluation
- MPNDocument15 pagesMPNShendi SuryanaPas encore d'évaluation
- 1 s2.0 S0304389407005407 MainDocument6 pages1 s2.0 S0304389407005407 MainFarah TalibPas encore d'évaluation
- 813 FullDocument11 pages813 Fullmicrobiolgy16Pas encore d'évaluation
- Flavonoids From and Their Antioxidant Activities: Vernonia AmygdalinaDocument4 pagesFlavonoids From and Their Antioxidant Activities: Vernonia AmygdalinatirahmPas encore d'évaluation
- Chlorella Distintos Fotoperiodos e IrradianciaDocument9 pagesChlorella Distintos Fotoperiodos e IrradianciaDelfina AlmeydaPas encore d'évaluation
- Denaturing Gradient Gel Electrophoresis of β-ThalassemiaDocument2 pagesDenaturing Gradient Gel Electrophoresis of β-Thalassemiau77Pas encore d'évaluation
- Chelex Protocol SLBDocument2 pagesChelex Protocol SLBu77Pas encore d'évaluation
- Microbial Diversity by DGGEDocument2 pagesMicrobial Diversity by DGGEu77Pas encore d'évaluation
- Methods For Studying CytotoxicityDocument2 pagesMethods For Studying Cytotoxicityu77Pas encore d'évaluation
- Dna Extraction Using Chelex ResinDocument7 pagesDna Extraction Using Chelex Resinu77Pas encore d'évaluation
- Chelex DNA Preparation Protocol ForDocument2 pagesChelex DNA Preparation Protocol Foru77Pas encore d'évaluation
- Chelating Resin - Product.informationDocument2 pagesChelating Resin - Product.informationu77Pas encore d'évaluation
- Chelex 100-Instruction ManualDocument14 pagesChelex 100-Instruction Manualu77Pas encore d'évaluation
- Syber Green Quantitative-PCRDocument60 pagesSyber Green Quantitative-PCRu77Pas encore d'évaluation
- Utility of The Housekeeping Genes 18S rRNA, B-Actin-04Document8 pagesUtility of The Housekeeping Genes 18S rRNA, B-Actin-04u77Pas encore d'évaluation
- Roche Primer DimerDocument4 pagesRoche Primer Dimeru77Pas encore d'évaluation
- Real Quant PCRDocument48 pagesReal Quant PCRu77Pas encore d'évaluation
- Quantitative RT-PCR Protocol (Sybr Green I)Document8 pagesQuantitative RT-PCR Protocol (Sybr Green I)u77Pas encore d'évaluation
- Real-Time Amplification On The Rotor-Gene: Overview of The Chemistries and OptimizationsDocument50 pagesReal-Time Amplification On The Rotor-Gene: Overview of The Chemistries and Optimizationsu77Pas encore d'évaluation
- Sybr Green PCR Master Mix and RT-PCR: ProtocolDocument64 pagesSybr Green PCR Master Mix and RT-PCR: Protocolu77Pas encore d'évaluation
- Real Time PCRDocument7 pagesReal Time PCRu77Pas encore d'évaluation
- Assay Formats For Use R-Time PCRDocument14 pagesAssay Formats For Use R-Time PCRu77Pas encore d'évaluation
- Real-Time PCRDocument5 pagesReal-Time PCRu77Pas encore d'évaluation
- UT Dallas Syllabus For Govt4396.002.08s Taught by Gregory Thielemann (Gregt)Document2 pagesUT Dallas Syllabus For Govt4396.002.08s Taught by Gregory Thielemann (Gregt)UT Dallas Provost's Technology GroupPas encore d'évaluation
- Chapter 13 (Automatic Transmission)Document26 pagesChapter 13 (Automatic Transmission)ZIBA KHADIBIPas encore d'évaluation
- Bolt Grade Markings and Strength ChartDocument2 pagesBolt Grade Markings and Strength ChartGregory GaschteffPas encore d'évaluation
- Caspar Hirschi - The Origins of Nationalism - An Alternative History From Ancient Rome To Early Modern Germany-Cambridge University Press (2012)Document255 pagesCaspar Hirschi - The Origins of Nationalism - An Alternative History From Ancient Rome To Early Modern Germany-Cambridge University Press (2012)Roc SolàPas encore d'évaluation
- QuexBook TutorialDocument14 pagesQuexBook TutorialJeffrey FarillasPas encore d'évaluation
- Catify To Satisfy - Simple Solutions For Creating A Cat-Friendly Home (PDFDrive)Document315 pagesCatify To Satisfy - Simple Solutions For Creating A Cat-Friendly Home (PDFDrive)Paz Libros100% (2)
- Pityriasis VersicolorDocument10 pagesPityriasis Versicolorketty putriPas encore d'évaluation
- Chapter 2Document13 pagesChapter 2Kumkumo Kussia KossaPas encore d'évaluation
- Building Services Planning Manual-2007Document122 pagesBuilding Services Planning Manual-2007razanmrm90% (10)
- Presentation About GyroscopesDocument24 pagesPresentation About GyroscopesgeenjunkmailPas encore d'évaluation
- Phytotherapy On CancerDocument21 pagesPhytotherapy On CancerSiddhendu Bhattacharjee100% (1)
- Julia Dito ResumeDocument3 pagesJulia Dito Resumeapi-253713289Pas encore d'évaluation
- PMP Assesment TestDocument17 pagesPMP Assesment Testwilliam collinsPas encore d'évaluation
- 16783Document51 pages16783uddinnadeemPas encore d'évaluation
- Sveba Dahlen - SRP240Document16 pagesSveba Dahlen - SRP240Paola MendozaPas encore d'évaluation
- SOL LogicDocument21 pagesSOL LogicJa RiveraPas encore d'évaluation
- Aleutia Solar Container ClassroomDocument67 pagesAleutia Solar Container ClassroomaleutiaPas encore d'évaluation
- John L. Selzer - Merit and Degree in Webster's - The Duchess of MalfiDocument12 pagesJohn L. Selzer - Merit and Degree in Webster's - The Duchess of MalfiDivya AggarwalPas encore d'évaluation
- Pg2022 ResultDocument86 pagesPg2022 ResultkapilPas encore d'évaluation
- 15 Day Detox ChallengeDocument84 pages15 Day Detox ChallengeDanii Supergirl Bailey100% (4)
- Introduction To Screenwriting UEADocument12 pagesIntroduction To Screenwriting UEAMartín SalasPas encore d'évaluation
- Chapter 23Document9 pagesChapter 23Javier Chuchullo TitoPas encore d'évaluation
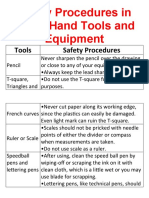
- Safety Procedures in Using Hand Tools and EquipmentDocument12 pagesSafety Procedures in Using Hand Tools and EquipmentJan IcejimenezPas encore d'évaluation
- Listen and Arrange The Sentences Based On What You Have Heard!Document3 pagesListen and Arrange The Sentences Based On What You Have Heard!Dewi Hauri Naura HaufanhazzaPas encore d'évaluation
- Tool Charts PDFDocument3 pagesTool Charts PDFtebengz100% (2)
- Sample Resume For Supply Chain Logistics PersonDocument2 pagesSample Resume For Supply Chain Logistics PersonAmmar AbbasPas encore d'évaluation
- HatfieldDocument33 pagesHatfieldAlex ForrestPas encore d'évaluation
- Shakespeare Sonnet EssayDocument3 pagesShakespeare Sonnet Essayapi-5058594660% (1)
- Applied Economics 2Document8 pagesApplied Economics 2Sayra HidalgoPas encore d'évaluation
- Friction: Ultiple Hoice UestionsDocument5 pagesFriction: Ultiple Hoice Uestionspk2varmaPas encore d'évaluation