Vous aimerez peut-être aussi
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- Structural Variations in Natural Fo Oh, and CL Apatites Jonn M. Hucnrs. Manvnr-R.Nx Cevrcnon. KNVRN D. Cnowr-BvDocument7 pagesStructural Variations in Natural Fo Oh, and CL Apatites Jonn M. Hucnrs. Manvnr-R.Nx Cevrcnon. KNVRN D. Cnowr-BvPoonam MondalPas encore d'évaluation
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- c99045 PDFDocument58 pagesc99045 PDFPoonam MondalPas encore d'évaluation
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
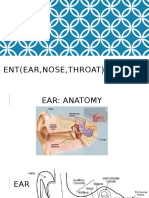
- Ent (Ear, Nose, Throat)Document14 pagesEnt (Ear, Nose, Throat)Poonam MondalPas encore d'évaluation
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- Environmental Legislation and Environmental ProtectionDocument26 pagesEnvironmental Legislation and Environmental ProtectionPoonam MondalPas encore d'évaluation
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Prostaglandins, Immunoglobulins: Prepared by Dr. Poonam MondalDocument10 pagesProstaglandins, Immunoglobulins: Prepared by Dr. Poonam MondalPoonam MondalPas encore d'évaluation
- Centre For Plant BiotechnologyDocument4 pagesCentre For Plant BiotechnologyPoonam MondalPas encore d'évaluation
- Blood and Circulation: Submitted By: Poonam B.T-M.T Biotech SEC: G77M2 Roll No: 37Document15 pagesBlood and Circulation: Submitted By: Poonam B.T-M.T Biotech SEC: G77M2 Roll No: 37Poonam MondalPas encore d'évaluation
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (400)
- Divine Intervention Biochem ReviewDocument2 pagesDivine Intervention Biochem Reviewmemmedoval752Pas encore d'évaluation
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Amino Acid MetabolismDocument4 pagesAmino Acid Metabolismclesleon100% (2)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- Methods in Protein EngineeringDocument5 pagesMethods in Protein EngineeringUmar EjazPas encore d'évaluation
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- Nomenclature and ClassificationDocument9 pagesNomenclature and ClassificationdkPas encore d'évaluation
- 3:5 BioDocument4 pages3:5 BioMaya FinklePas encore d'évaluation
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- 1 MCQ Biochemistry For - Paramedical - Students PDFDocument78 pages1 MCQ Biochemistry For - Paramedical - Students PDFmuhammad shuaib86% (7)
- Types of RNA-Worksheet Omar IbrahimDocument5 pagesTypes of RNA-Worksheet Omar IbrahimOmar Ibrahim LPas encore d'évaluation
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- Homology Modelling and AutodockDocument25 pagesHomology Modelling and AutodockGsbsggsbsjysbsnshsbPas encore d'évaluation
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (345)
- BIO156 Enzyme Lab-Effect of Temperature On Enzyme Activity-F21Document8 pagesBIO156 Enzyme Lab-Effect of Temperature On Enzyme Activity-F21Shiloh FrederickPas encore d'évaluation
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (74)
- Pharmaceutical Biochemistry (PACOP RED)Document20 pagesPharmaceutical Biochemistry (PACOP RED)Fred0% (1)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- Post-Translational Modifications of Proteins: Acetylcholinesterase As A Model SystemDocument17 pagesPost-Translational Modifications of Proteins: Acetylcholinesterase As A Model SystemSergio AraujoPas encore d'évaluation
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- BiuretDocument10 pagesBiuretIndrie DwiraandaPas encore d'évaluation
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The Three-Dimensional Structure of Proteins: Protein Structure and FunctionDocument26 pagesThe Three-Dimensional Structure of Proteins: Protein Structure and Function김경환Pas encore d'évaluation
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- John K. Mathai, Yanhong Liu and Hans H. SteinDocument10 pagesJohn K. Mathai, Yanhong Liu and Hans H. SteinDébora Giaretta ZattaPas encore d'évaluation
- Biochemistry Lecture 15 Biological Membranes - 1Document27 pagesBiochemistry Lecture 15 Biological Membranes - 1Professor Rakesh Sharma Biochemistry Lectures100% (1)
- Amino Acids, Classifications, Properties and FunctionsDocument22 pagesAmino Acids, Classifications, Properties and FunctionsChe ChePas encore d'évaluation
- S2C - Activity Sheet - Activity 2 1 2Document2 pagesS2C - Activity Sheet - Activity 2 1 2lucerocatapanggwen.03Pas encore d'évaluation
- Billesblle 2023 Structural Basis of Odorant RecogniDocument30 pagesBillesblle 2023 Structural Basis of Odorant RecogniAgnes_Agnes_AgnesPas encore d'évaluation
- Amino Acids in Animal NutritionDocument515 pagesAmino Acids in Animal NutritionLuciana EmanuellePas encore d'évaluation
- Enzymes: Anushree ChaturvediDocument39 pagesEnzymes: Anushree ChaturvediDeepal ShahPas encore d'évaluation
- Western Blotting of Phospho-Proteins ProtocolDocument1 pageWestern Blotting of Phospho-Proteins ProtocolGabriela VenturiniPas encore d'évaluation
- Titin ProteinDocument40 pagesTitin ProteinAlexanderPas encore d'évaluation
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- 5 Enzymes 1Document11 pages5 Enzymes 1diahema50% (2)
- CHAPTER 3: Genetic Control of Protein Synthesis, Cell Function, and Cell ReproductionDocument4 pagesCHAPTER 3: Genetic Control of Protein Synthesis, Cell Function, and Cell ReproductionMan DejeloPas encore d'évaluation
- Protein PDFDocument36 pagesProtein PDF2d HoehoePas encore d'évaluation
- Introduction To BiomoleculesDocument5 pagesIntroduction To BiomoleculesGhaiiPas encore d'évaluation
- Solutions To 7.012 Problem Set 1Document4 pagesSolutions To 7.012 Problem Set 1attiquePas encore d'évaluation
- Dna - CotDocument39 pagesDna - CotJcob BangcayaPas encore d'évaluation
- When Insulin Molecules Are Formed They Fold Into A Specific ShapeDocument6 pagesWhen Insulin Molecules Are Formed They Fold Into A Specific Shapenaruto1221Pas encore d'évaluation
- Quiz 1 - BIOL 420 U - Advanced Cell and Molecular BiologyDocument6 pagesQuiz 1 - BIOL 420 U - Advanced Cell and Molecular BiologyCumPas encore d'évaluation
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)