Vous aimerez peut-être aussi
- Conferences: and ReviewsDocument10 pagesConferences: and ReviewsGeorgarasgtPas encore d'évaluation
- HUGO Gene Nomenclature Committee (HGNC) Recommendations For The Designation of Gene FusionsDocument4 pagesHUGO Gene Nomenclature Committee (HGNC) Recommendations For The Designation of Gene Fusionsdaniagurendon92Pas encore d'évaluation
- Oncogenes and Proto-Oncogenes: of ofDocument5 pagesOncogenes and Proto-Oncogenes: of ofSilvia SoarePas encore d'évaluation
- TMP 3Document3 pagesTMP 3FrontiersPas encore d'évaluation
- Genetic Alterations in The Adenoma-Carcinoma Sequence: Kathleen R. ChoDocument5 pagesGenetic Alterations in The Adenoma-Carcinoma Sequence: Kathleen R. ChoSantiago AldayPas encore d'évaluation
- Meiosis PDFDocument54 pagesMeiosis PDFYusrahanna daulayPas encore d'évaluation
- Medical Progress: Review ArticlesDocument11 pagesMedical Progress: Review ArticlesEllie satrianiPas encore d'évaluation
- CancerDocument20 pagesCancerManisha DasPas encore d'évaluation
- Three Dimensional Genome Architecture Players and MechanismsDocument13 pagesThree Dimensional Genome Architecture Players and MechanismsNatalio Vazquez CruzPas encore d'évaluation
- Describe The Molecular Basis Underlying Haematological MalignancyDocument3 pagesDescribe The Molecular Basis Underlying Haematological MalignancyAlison HinesPas encore d'évaluation
- Globin Synthesis PDFDocument6 pagesGlobin Synthesis PDFmanoj_rkl_07100% (1)
- 0039-Bh-Bio-T-19 (Muhammad Ahsan)Document9 pages0039-Bh-Bio-T-19 (Muhammad Ahsan)خاک نشینPas encore d'évaluation
- Survey and Summary How Does DNA Break During Chromosomal Translocations?Document13 pagesSurvey and Summary How Does DNA Break During Chromosomal Translocations?Daniel ALPas encore d'évaluation
- CML MiceDocument14 pagesCML MiceKiran MandapatiPas encore d'évaluation
- Mechanisms of Eukaryotic DNA RecombinationD'EverandMechanisms of Eukaryotic DNA RecombinationMax E GottesmanPas encore d'évaluation
- Biochemistry of Sickle Cell Disease and Related HaemoglobinopathiesDocument9 pagesBiochemistry of Sickle Cell Disease and Related HaemoglobinopathiesClement BewajiPas encore d'évaluation
- Hematology. 1Document10 pagesHematology. 1Krismas Eka SaputraPas encore d'évaluation
- Case StudiesDocument14 pagesCase StudiesS NairPas encore d'évaluation
- TrombopoiesisDocument8 pagesTrombopoiesisKie SanPas encore d'évaluation
- Presentation OncogenesDocument54 pagesPresentation Oncogenesyash mathurPas encore d'évaluation
- Cell Death Induced by DexamethasoneDocument12 pagesCell Death Induced by DexamethasoneJesus_Arraiz_14Pas encore d'évaluation
- How Homology' Entered Genetics: OutlookDocument3 pagesHow Homology' Entered Genetics: OutlookSebas CirignoliPas encore d'évaluation
- Identification of Recurring Tumor Specific Somatic Mutations in AML by Transcriptome Seq Greif2011Document7 pagesIdentification of Recurring Tumor Specific Somatic Mutations in AML by Transcriptome Seq Greif2011mrkhprojectPas encore d'évaluation
- A Novel Somatic K-Ras Mutation in Juvenile Myelomonocytic LeukemiaDocument2 pagesA Novel Somatic K-Ras Mutation in Juvenile Myelomonocytic LeukemiamahanteshPas encore d'évaluation
- 2014 Torres (Nat. Comm) - Engineering Human Tumour-Associated Chromosomal Translocations With The RNA-guided CRISPR-Cas9 SystemDocument8 pages2014 Torres (Nat. Comm) - Engineering Human Tumour-Associated Chromosomal Translocations With The RNA-guided CRISPR-Cas9 SystemJenny ChenPas encore d'évaluation
- Role of Break Cluster Region (BCR) - Abelson Murine Leukimia (Abl) Examination in Chronic Myelogenous Leukemia (CML)Document4 pagesRole of Break Cluster Region (BCR) - Abelson Murine Leukimia (Abl) Examination in Chronic Myelogenous Leukemia (CML)AdjenkRamadhaniPas encore d'évaluation
- Colwrt 161 Mini-Lit-ReviewDocument18 pagesColwrt 161 Mini-Lit-Reviewapi-626218608Pas encore d'évaluation
- Human Chromosomes: An Illustrated Introduction to Human CytogeneticsD'EverandHuman Chromosomes: An Illustrated Introduction to Human CytogeneticsÉvaluation : 5 sur 5 étoiles5/5 (1)
- 2019 CHROMOTRYPSIS UpdateDocument15 pages2019 CHROMOTRYPSIS UpdatemaomaochongPas encore d'évaluation
- Cell Sept 2011Document201 pagesCell Sept 2011mekkararameshPas encore d'évaluation
- Chromosome Translocations and CancerDocument5 pagesChromosome Translocations and CancervictoryPas encore d'évaluation
- 976 FullDocument7 pages976 FullFirda PotterPas encore d'évaluation
- Gibas Et Al. - Nonrandom Chromosomal Changes in TCC of The BladderDocument9 pagesGibas Et Al. - Nonrandom Chromosomal Changes in TCC of The BladderyuenkeithPas encore d'évaluation
- 1 s2.0 S1535947621001584 MainDocument28 pages1 s2.0 S1535947621001584 MainBen DresimPas encore d'évaluation
- 1a - Three Dimensional Genome Architecture Players and MechanismsDocument13 pages1a - Three Dimensional Genome Architecture Players and MechanismsStephanePas encore d'évaluation
- 1 s2.0 S0955067416300333 Main 2Document8 pages1 s2.0 S0955067416300333 Main 2maomaochongPas encore d'évaluation
- Mutations in The APC Tumor Suppressor Gene Cause Chromosomal InstabilityDocument10 pagesMutations in The APC Tumor Suppressor Gene Cause Chromosomal InstabilityRBrian KielmanPas encore d'évaluation
- tmp8556 TMPDocument26 pagestmp8556 TMPFrontiersPas encore d'évaluation
- Boyd 1988Document10 pagesBoyd 1988YEKDA KARACANPas encore d'évaluation
- 트랜스포존Document12 pages트랜스포존100 568 최재훈Pas encore d'évaluation
- GKT 1005Document13 pagesGKT 1005SairaBurciagaPas encore d'évaluation
- Tumor ProgressionDocument7 pagesTumor ProgressionmineresearchPas encore d'évaluation
- Major Project Proposal - BIODocument10 pagesMajor Project Proposal - BIOsappy490Pas encore d'évaluation
- Cytogenetic Analysis in The Diagnosis of Acute Leukemia: Sverre Heim, FelixDocument9 pagesCytogenetic Analysis in The Diagnosis of Acute Leukemia: Sverre Heim, FelixEnas KharbotlyPas encore d'évaluation
- Chronic Myelogenous LeukemiaDocument25 pagesChronic Myelogenous LeukemiaarieftamaPas encore d'évaluation
- ZET 403 Cyto Extra Nuclear InheritenceDocument15 pagesZET 403 Cyto Extra Nuclear InheritenceblazeaniikPas encore d'évaluation
- If You Understand How Proteins WorkDocument4 pagesIf You Understand How Proteins WorkMah Song WeiPas encore d'évaluation
- Ass and CharactDocument6 pagesAss and CharactAsmaa Nasr-El-DinePas encore d'évaluation
- Spur Beck 2004sDocument18 pagesSpur Beck 2004snvp16Pas encore d'évaluation
- Articol Clonare CFTRDocument1 pageArticol Clonare CFTRPancu CristianaPas encore d'évaluation
- Review Article: Apoptosis in YeastsDocument6 pagesReview Article: Apoptosis in YeastsShreya AhujaPas encore d'évaluation
- Molecularmarkersfor Colorectalcancer: Moriah Wright,, Jenifer S. Beaty,, Charles A. TernentDocument19 pagesMolecularmarkersfor Colorectalcancer: Moriah Wright,, Jenifer S. Beaty,, Charles A. TernentFernando Castro EchavezPas encore d'évaluation
- HOMOLOGOUS AND NON HOMOLOGOUS RECOMBINATION by Preeti Saini 21mslsmm06Document7 pagesHOMOLOGOUS AND NON HOMOLOGOUS RECOMBINATION by Preeti Saini 21mslsmm06Preeti SainiPas encore d'évaluation
- Cancer Res 1990 Wainscoat 1355 60Document7 pagesCancer Res 1990 Wainscoat 1355 60Shahab Ud DinPas encore d'évaluation
- p53 in Leukernogenesis of Chronic Myeloid Leukemia: Francesco Lanza, A?' SucaiDocument8 pagesp53 in Leukernogenesis of Chronic Myeloid Leukemia: Francesco Lanza, A?' SucaiEduardo Martín Ramírez López.Pas encore d'évaluation
- Ch. 9 Cell CycleDocument2 pagesCh. 9 Cell Cycleapi-290697718Pas encore d'évaluation
- Mycobacterial ChaperoninsDocument6 pagesMycobacterial Chaperoninscamilo.colacoPas encore d'évaluation
- Protein Diversity 1Document4 pagesProtein Diversity 1Lalitha RajeshPas encore d'évaluation
- Order and Stochastic Dynamics in Drosophila Planar Cell PolarityDocument10 pagesOrder and Stochastic Dynamics in Drosophila Planar Cell PolarityLuu XiaoPas encore d'évaluation
- 3 1 and 3 2Document28 pages3 1 and 3 2api-235355872Pas encore d'évaluation
- Journal of Visualized ExperimentsDocument4 pagesJournal of Visualized ExperimentsNatasa BajalovicPas encore d'évaluation
- LEC9 InstabilityDocument40 pagesLEC9 InstabilityNatasa BajalovicPas encore d'évaluation
- Alchajmerova BolestDocument23 pagesAlchajmerova BolestNatasa BajalovicPas encore d'évaluation
- Cancer General and TherapyDocument62 pagesCancer General and TherapyNatasa BajalovicPas encore d'évaluation
- Introductory Q Bank (Pathology MCQ)Document9 pagesIntroductory Q Bank (Pathology MCQ)EZHIL N100% (1)
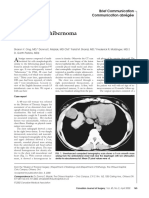
- Intrathoracic Hibernoma: Brief Communication Communication AbrégéeDocument2 pagesIntrathoracic Hibernoma: Brief Communication Communication AbrégéeM Ali AdrianPas encore d'évaluation
- Cancer Healing MantraDocument2 pagesCancer Healing MantraAtul SinghPas encore d'évaluation
- Baneji PDFDocument6 pagesBaneji PDFSuraj YadavPas encore d'évaluation
- The FRCPath Exam - Paul BennettDocument130 pagesThe FRCPath Exam - Paul BennettVictor ChanPas encore d'évaluation
- Testicular TumorsDocument42 pagesTesticular TumorsarhamPas encore d'évaluation
- Radio DNB Q PapersDocument44 pagesRadio DNB Q PapersRakhi SuryawanshiPas encore d'évaluation
- Soalan Pathology 2Document347 pagesSoalan Pathology 2muhammadridhwanPas encore d'évaluation
- Tumors of The Fetus and Infant-An Atlas PDFDocument435 pagesTumors of The Fetus and Infant-An Atlas PDFAntohi Catalin100% (1)
- Anticancer and Antiblastomic - DrugsDocument29 pagesAnticancer and Antiblastomic - DrugsmsrchesustPas encore d'évaluation
- Detection of Brain Tumors Using CNNDocument3 pagesDetection of Brain Tumors Using CNNKshatriya KulkesPas encore d'évaluation
- Oxo AQAGCSE B7 ws01 XxaannDocument2 pagesOxo AQAGCSE B7 ws01 XxaannParam BhimaniPas encore d'évaluation
- Ringkasan Chapter Orbita Buku GrangerDocument35 pagesRingkasan Chapter Orbita Buku GrangerIrni Dwi Aprianty IbrahimPas encore d'évaluation
- Frozen Section Library Gynecologic Pathology Intraoperative Consultation 2012 PGDocument253 pagesFrozen Section Library Gynecologic Pathology Intraoperative Consultation 2012 PGOlteanu Dragos-Nicolae100% (1)
- Atlas Poultry Diseases (Picture Book) - Scanned BookDocument107 pagesAtlas Poultry Diseases (Picture Book) - Scanned BookSuraj_Subedi92% (37)
- Ca MammaeDocument18 pagesCa MammaeRizmaMudzalifahPas encore d'évaluation
- Annsurg00602 0014Document7 pagesAnnsurg00602 0014MogiMediawanPas encore d'évaluation
- Wilms Tumor: Imaging of Pediatric Renal MassesDocument35 pagesWilms Tumor: Imaging of Pediatric Renal MassesEnny AndrianiPas encore d'évaluation
- Patho QuestDocument4 pagesPatho QuestRosel Ann BontiaPas encore d'évaluation
- Human Cancer CellsDocument9 pagesHuman Cancer Cellssai collegePas encore d'évaluation
- Management of An Adnexal Mass UpToDateDocument27 pagesManagement of An Adnexal Mass UpToDateAMGY82Pas encore d'évaluation
- The Cell Cycle and Cancer 2011Document28 pagesThe Cell Cycle and Cancer 2011Rajesh BalakrishnanPas encore d'évaluation
- Cytodiagnosis in DermatologyDocument5 pagesCytodiagnosis in DermatologyDipti MathiasPas encore d'évaluation
- CRCDocument5 pagesCRCanon_619577898Pas encore d'évaluation
- HistopathDocument37 pagesHistopathFait HeePas encore d'évaluation
- AngiofibromaDocument7 pagesAngiofibromaPandu DiputraPas encore d'évaluation
- Malignant Soft Tissue TumorsDocument21 pagesMalignant Soft Tissue TumorsEva GustianiPas encore d'évaluation
- Skin Tumor AgusDocument54 pagesSkin Tumor AgusHafida AuliaristaPas encore d'évaluation
- PET CT Case Based ApproachDocument316 pagesPET CT Case Based ApproachAzmal Kabir SarkerPas encore d'évaluation
- Tumour Suppressor Genes CLS 712 2022-2023Document35 pagesTumour Suppressor Genes CLS 712 2022-2023Heba LahhamPas encore d'évaluation