Vous aimerez peut-être aussi
- Aldehidos y CetonasDocument32 pagesAldehidos y Cetonasalfayomega48388% (16)
- 16 - Apola OfunDocument92 pages16 - Apola Ofunrota204100% (1)
- La Estrategia Ateisante de Antonio Gramsci - Alfredo Sáenz, S. J.Document14 pagesLa Estrategia Ateisante de Antonio Gramsci - Alfredo Sáenz, S. J.moltenpaper100% (1)
- Winny-Farma 2 Sox UnitedDocument208 pagesWinny-Farma 2 Sox Unitedarchivos primeroPas encore d'évaluation
- Vamos Creciendo, Vamos Cambiando. TeoriaDocument2 pagesVamos Creciendo, Vamos Cambiando. TeoriaEduardo Cardenas Campos0% (1)
- TetradotoxinaDocument15 pagesTetradotoxinaVíctor AlcántarPas encore d'évaluation
- Bioquimica DigestivoDocument90 pagesBioquimica DigestivoAlan Mendoza100% (1)
- Alambres en OrtodonciaDocument102 pagesAlambres en OrtodonciaNancy Chaparro100% (1)
- AnaRojas TI Auditoria Estados FinancierosDocument7 pagesAnaRojas TI Auditoria Estados FinancierosAguilas Agustinas Voley100% (1)
- Clase 6 MetabolismoDocument51 pagesClase 6 MetabolismoFrancesco Andrés Placencia SquadritoPas encore d'évaluation
- Sistema de ComplementoDocument6 pagesSistema de ComplementoPhanny Ni SandovalPas encore d'évaluation
- Sistema Inmune 2018 Tipeo 2Document34 pagesSistema Inmune 2018 Tipeo 2Belen Hernandez AguilarPas encore d'évaluation
- Principios Generales Tratamiento AntimicrobianoDocument10 pagesPrincipios Generales Tratamiento AntimicrobianoDr. MSc. Byron Núñez FreilePas encore d'évaluation
- Método de La AutoobservaciónDocument4 pagesMétodo de La AutoobservaciónRodrigo Vargas PérezPas encore d'évaluation
- Directorio Facultad Medicina UNAMDocument74 pagesDirectorio Facultad Medicina UNAMCarlos Ivan Chavesta RimapaPas encore d'évaluation
- Lectura de InmunidadDocument8 pagesLectura de InmunidadKatherine Milagros Apcho VeraPas encore d'évaluation
- Ciclo celular mitosis meiosisDocument8 pagesCiclo celular mitosis meiosisAle Aguirre LopezPas encore d'évaluation
- Diureticos-Isaza Ocr PDFDocument10 pagesDiureticos-Isaza Ocr PDFSantiago Figueroa TorresPas encore d'évaluation
- Sistema de Complemento y Sistema Inmune de Mucosas - InflamacionDocument32 pagesSistema de Complemento y Sistema Inmune de Mucosas - InflamacionSantiago CortesPas encore d'évaluation
- Histología Del Sistema InmuneDocument9 pagesHistología Del Sistema InmuneJosé Luis Juárez VegaPas encore d'évaluation
- HematopoyesisDocument13 pagesHematopoyesisLEO 999Pas encore d'évaluation
- Quimioterapia antibacteriana: mecanismos y clasificaciónDocument13 pagesQuimioterapia antibacteriana: mecanismos y clasificaciónValentina GutierrezPas encore d'évaluation
- LisosomasDocument14 pagesLisosomasAndrea0% (1)
- Extraccion de Adn A Partir de Sangre Periferica 1Document4 pagesExtraccion de Adn A Partir de Sangre Periferica 1PamelaaGaytan100% (1)
- RenalDocument6 pagesRenalClaudia Carbajal Castro60% (5)
- Inflamación y Reparación TisularDocument11 pagesInflamación y Reparación TisularBetty CarreñoPas encore d'évaluation
- Barreras AnatómicasDocument7 pagesBarreras AnatómicasFay CerónPas encore d'évaluation
- Formula LeucocitariaDocument5 pagesFormula LeucocitariaJesus Loreto RangelPas encore d'évaluation
- EXTRACCIÓN DE ADN (Ácido DesoxirribonucleicoDocument2 pagesEXTRACCIÓN DE ADN (Ácido DesoxirribonucleicoEric Bertel SalazarPas encore d'évaluation
- Patologias Por Deficencia de EnzimasDocument3 pagesPatologias Por Deficencia de EnzimasmaalhuesPas encore d'évaluation
- Practica #1Document8 pagesPractica #1Carlos AnayaPas encore d'évaluation
- Células Del Sistema InmunológicoDocument7 pagesCélulas Del Sistema InmunológicoDamian Junior G. LassondePas encore d'évaluation
- Presentacion Enzimas 1 Generalidades, Nomenclatura y ClasificacionDocument58 pagesPresentacion Enzimas 1 Generalidades, Nomenclatura y ClasificacionMariana Maya GuerreroPas encore d'évaluation
- Biología celular y enzimasDocument23 pagesBiología celular y enzimasDario Neri CortezPas encore d'évaluation
- Mecanismos de acción antimicrobianosDocument78 pagesMecanismos de acción antimicrobianosJOSE ANGEL LAZARO BARRERAPas encore d'évaluation
- Bacterias PrototrofasDocument1 pageBacterias PrototrofasJosé David Ronquillo CandoPas encore d'évaluation
- Estado actual de la transfusión de hemocomponentesDocument58 pagesEstado actual de la transfusión de hemocomponentesCarlos BuitragoPas encore d'évaluation
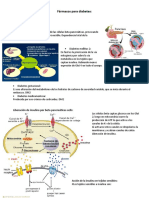
- Fármacos para la diabetesDocument6 pagesFármacos para la diabetesCRISTÓBAL IGNACIO AROSPas encore d'évaluation
- Deficiencias de Las Enzimas Del EritrocitosDocument7 pagesDeficiencias de Las Enzimas Del EritrocitosAngélica RojasPas encore d'évaluation
- Genetica BacterianaDocument43 pagesGenetica BacterianaVickyGaliciaPas encore d'évaluation
- Síntesis ATP mitocondria: gradiente protones produce ATPDocument2 pagesSíntesis ATP mitocondria: gradiente protones produce ATPDaniel Steven Muñoz CastilloPas encore d'évaluation
- Cap 7 Enzimas Mecanismo de AcciónDocument6 pagesCap 7 Enzimas Mecanismo de AcciónGmo8 CPas encore d'évaluation
- El Sistema Del ComplementoDocument7 pagesEl Sistema Del ComplementojesusPas encore d'évaluation
- LPS y Receptores Toll (MAOG)Document13 pagesLPS y Receptores Toll (MAOG)miguel angel100% (3)
- Acidos Nucleicos y MutacionesDocument23 pagesAcidos Nucleicos y Mutacionesluiferalxeliu1Pas encore d'évaluation
- Daño Celular y Patogénesis ViralDocument16 pagesDaño Celular y Patogénesis ViralAlejandro Villagrana AlvarezPas encore d'évaluation
- Conversatorio 13-2018 IDocument2 pagesConversatorio 13-2018 IAtenas VargasPas encore d'évaluation
- Cascada de La CoagulaciónDocument3 pagesCascada de La CoagulaciónTosCof100% (2)
- Señalización CelularDocument12 pagesSeñalización Celularisaac jimenez mendozaPas encore d'évaluation
- Taller EnzimaDocument7 pagesTaller EnzimaHanner ValegaPas encore d'évaluation
- Ciclo de KrebsDocument21 pagesCiclo de KrebsAndresPas encore d'évaluation
- ENZIMOLOGÍA CLÍNICA SeminarioDocument22 pagesENZIMOLOGÍA CLÍNICA SeminarioDante TorresPas encore d'évaluation
- MICROAMBIENTE MEDULAR REGULA CELULAS MADREDocument11 pagesMICROAMBIENTE MEDULAR REGULA CELULAS MADREGido Cliner Quispe FernandezPas encore d'évaluation
- Toma de MuestrasDocument52 pagesToma de MuestrasShannen Mileth VizcaínoPas encore d'évaluation
- Funciones Del AmpcDocument3 pagesFunciones Del AmpcRaulPas encore d'évaluation
- Introducción A La FarmacologíaDocument17 pagesIntroducción A La FarmacologíaCamila Briceño GómezPas encore d'évaluation
- Sistema inmunitario: definición, componentes y funcionesDocument17 pagesSistema inmunitario: definición, componentes y funcionesCarlos Zanns CarpioPas encore d'évaluation
- División Celular Sin ControlDocument7 pagesDivisión Celular Sin ControlLizbeth AlegriaPas encore d'évaluation
- Desgrabacion: Sistema de ComplementoDocument12 pagesDesgrabacion: Sistema de ComplementoVeronica MendozaPas encore d'évaluation
- Fisiologia LeucocitariaDocument7 pagesFisiologia LeucocitariaJaviera Alejandra Montoya AcevedoPas encore d'évaluation
- Cortes histológicos tejidos biológicosDocument6 pagesCortes histológicos tejidos biológicosSalvador GallardoPas encore d'évaluation
- Exámenes Macroscópicos, Microscópicos y QuimicosDocument4 pagesExámenes Macroscópicos, Microscópicos y QuimicosLuisaRodriguezPas encore d'évaluation
- Generalidades Del NúcleoDocument5 pagesGeneralidades Del NúcleoCarlo CepulvedaPas encore d'évaluation
- Bioquímica: Lípidos y membranasDocument63 pagesBioquímica: Lípidos y membranasChuritos Nena VeintimillaPas encore d'évaluation
- Biocatalizadores PDFDocument93 pagesBiocatalizadores PDFJuan Camilo Gonzalez100% (1)
- (Sem) Bioquimica - EnzimasDocument26 pages(Sem) Bioquimica - EnzimasUSMP FN ARCHIVOS100% (3)
- Enzimas, Cinetica y RegulaciónDocument55 pagesEnzimas, Cinetica y RegulaciónAlas25Pas encore d'évaluation
- Esterificación TeoriaDocument7 pagesEsterificación TeoriaJossi Maggali Soriano PomayaPas encore d'évaluation
- Benceno ToluenoDocument1 pageBenceno ToluenoJossi Maggali Soriano PomayaPas encore d'évaluation
- Problemas de MoliendaDocument5 pagesProblemas de MoliendaJossi Maggali Soriano PomayaPas encore d'évaluation
- Cálculos y Resultados InstrumentacionDocument2 pagesCálculos y Resultados InstrumentacionJossi Maggali Soriano PomayaPas encore d'évaluation
- Equilibrio Liquido - ImprimirDocument15 pagesEquilibrio Liquido - ImprimirJossi Maggali Soriano PomayaPas encore d'évaluation
- Cálculos y Resultados InstrumentacionDocument2 pagesCálculos y Resultados InstrumentacionJossi Maggali Soriano PomayaPas encore d'évaluation
- Equilibrio Liquido - ImprimirDocument15 pagesEquilibrio Liquido - ImprimirJossi Maggali Soriano PomayaPas encore d'évaluation
- Problemas de MoliendaDocument5 pagesProblemas de MoliendaJossi Maggali Soriano PomayaPas encore d'évaluation
- Cálculos y Resultados InstrumentacionDocument2 pagesCálculos y Resultados InstrumentacionJossi Maggali Soriano PomayaPas encore d'évaluation
- Bicarbonatos y ClorurosDocument3 pagesBicarbonatos y ClorurosJossi Maggali Soriano PomayaPas encore d'évaluation
- Control Trabajo D EapliDocument33 pagesControl Trabajo D EapliJossi Maggali Soriano PomayaPas encore d'évaluation
- Propuesta Detratamiento de Gases ContaminantesDocument24 pagesPropuesta Detratamiento de Gases ContaminantesJossi Maggali Soriano PomayaPas encore d'évaluation
- Huella Ecologica, Naturaleza y BiodiversidadDocument10 pagesHuella Ecologica, Naturaleza y BiodiversidadJossi Maggali Soriano PomayaPas encore d'évaluation
- Contaminacion Del AireDocument1 pageContaminacion Del AireJossi Maggali Soriano PomayaPas encore d'évaluation
- Oferta y DemandaDocument13 pagesOferta y DemandaJossi Maggali Soriano PomayaPas encore d'évaluation
- Agua, Aire y SueloDocument26 pagesAgua, Aire y SueloJossi Maggali Soriano PomayaPas encore d'évaluation
- CALCIODocument2 pagesCALCIOJossi Maggali Soriano PomayaPas encore d'évaluation
- Clasificación ciencias 40Document2 pagesClasificación ciencias 40Jossi Maggali Soriano Pomaya100% (1)
- Soteriología 1Document5 pagesSoteriología 1Elias PontePas encore d'évaluation
- Universidad Nacional de Piura Facultad de Ingenieria Civil Mecanica de Suelos 1 - Examen Sustitutorio - 2018.1 IndicacionesDocument6 pagesUniversidad Nacional de Piura Facultad de Ingenieria Civil Mecanica de Suelos 1 - Examen Sustitutorio - 2018.1 Indicacionesclaudia valladares carrascoPas encore d'évaluation
- Soleal Fy 0267 007 1trim 2018 ESDocument25 pagesSoleal Fy 0267 007 1trim 2018 ESJuandaCabreraCoboPas encore d'évaluation
- Beneficios Anglo Salud (-65)Document3 pagesBeneficios Anglo Salud (-65)GCabrejoPas encore d'évaluation
- Introducción Al Caso DoraDocument7 pagesIntroducción Al Caso DoraAlejandra LeonPas encore d'évaluation
- Prelab 3 - Cinetica de Decoloracion FenolftaleinaDocument12 pagesPrelab 3 - Cinetica de Decoloracion FenolftaleinaBenjamin Quena CastroPas encore d'évaluation
- Introducción Historia de La Fitopatología 2017Document36 pagesIntroducción Historia de La Fitopatología 2017Lophophora930% (1)
- Calentador de Vapor DirectoDocument7 pagesCalentador de Vapor DirectoJORDYPas encore d'évaluation
- 4° Grado - Educación Primaria: Razonamiento VerbalDocument142 pages4° Grado - Educación Primaria: Razonamiento VerbalLesly GoicocheaPas encore d'évaluation
- Análisis Configuracional de Compuestos OrgánicosDocument16 pagesAnálisis Configuracional de Compuestos OrgánicosAgustín Isla0% (1)
- Cap 1 - Elementos Escenciales Del Capital HumanoDocument105 pagesCap 1 - Elementos Escenciales Del Capital HumanoRamon RodriguezPas encore d'évaluation
- IntructivoProcesos Gerenciales IngetecDocument95 pagesIntructivoProcesos Gerenciales Ingetecnathalia giraldoPas encore d'évaluation
- Tema 4: Memoria Interna. Tipos. Direccionamiento. Características y FuncionesDocument8 pagesTema 4: Memoria Interna. Tipos. Direccionamiento. Características y FuncionesMartin RucobaPas encore d'évaluation
- Modelo Registro Productos FitosanitariosDocument5 pagesModelo Registro Productos FitosanitariosCultivo del AlmendroPas encore d'évaluation
- Ciencias 7Document252 pagesCiencias 7Luis MendozaPas encore d'évaluation
- Cementaciones Con TFDocument39 pagesCementaciones Con TFjomarlucaPas encore d'évaluation
- Hay Que Ajustar Primero La Dureza Cálcica o La Alcalinidad en Una PiscinaDocument2 pagesHay Que Ajustar Primero La Dureza Cálcica o La Alcalinidad en Una PiscinaNilsPas encore d'évaluation
- Tesis Arquitectura Centro Deportivo PDFDocument106 pagesTesis Arquitectura Centro Deportivo PDFJonathan HernandezPas encore d'évaluation
- Visita A La Cadena SER.Document23 pagesVisita A La Cadena SER.Mara González GómezPas encore d'évaluation
- La Psicofisiológica y Su Objeto de EstudioDocument5 pagesLa Psicofisiológica y Su Objeto de EstudioGerick CalderónPas encore d'évaluation
- Producto AcademicoDocument31 pagesProducto AcademicoAnto PMontoyaPas encore d'évaluation
- Matriz de Riesgo ESCALONA 2022.Document7 pagesMatriz de Riesgo ESCALONA 2022.Bernardo MenesesPas encore d'évaluation
- Tarjeta Emergencia Varsol PolitecnicoDocument4 pagesTarjeta Emergencia Varsol PolitecnicoSofiaPas encore d'évaluation
- Lab Transistor BJTDocument11 pagesLab Transistor BJTalbertwillyPas encore d'évaluation