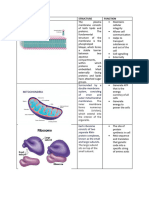
Vous aimerez peut-être aussi
- Cell Molecular Composition of CellDocument72 pagesCell Molecular Composition of Celljomel rondinaPas encore d'évaluation
- Eukaryotic and Prokaryotic CellDocument11 pagesEukaryotic and Prokaryotic CellAvyanna MillerPas encore d'évaluation
- CellDocument5 pagesCellhkrybmzxfbxbwnpfhnPas encore d'évaluation
- Common Features.: Omnis Cellula e CellulaDocument7 pagesCommon Features.: Omnis Cellula e CellulaTherese Jan MangussadPas encore d'évaluation
- Human Physiology Cell Structure Function I Lecture 31.july - .2012Document21 pagesHuman Physiology Cell Structure Function I Lecture 31.july - .2012Aakash ReddyPas encore d'évaluation
- Geral Aula #2b - Bioquimica e Biologia CelularDocument39 pagesGeral Aula #2b - Bioquimica e Biologia CelularsusanajpPas encore d'évaluation
- Chem123 - PrelimsDocument20 pagesChem123 - Prelimslily of the valleyPas encore d'évaluation
- Review of Cell Biology Concepts: Differences Between Prokaryotic Vs Eukaryotic CellsDocument25 pagesReview of Cell Biology Concepts: Differences Between Prokaryotic Vs Eukaryotic Cellsevil twinPas encore d'évaluation
- Cellular Structure: A Guide to the Organelles and Components of the CellDocument24 pagesCellular Structure: A Guide to the Organelles and Components of the CellTyra Joyce RevadaviaPas encore d'évaluation
- Group 2. Presentation - Cell 1Document57 pagesGroup 2. Presentation - Cell 1Thông LêPas encore d'évaluation
- 2 A - The-CellDocument18 pages2 A - The-CellSharmain Pranne Dulnuan NgipolPas encore d'évaluation
- 2.2 Tructures and Functions: Cell Membrane and OrganellesDocument8 pages2.2 Tructures and Functions: Cell Membrane and Organelles2023820042Pas encore d'évaluation
- 2.1 Cell Structure and FunctionDocument56 pages2.1 Cell Structure and Functionazrixone54Pas encore d'évaluation
- CHEM 142_N-BIOCHEMISTRY FOR NURSING CHAPTER 1Document10 pagesCHEM 142_N-BIOCHEMISTRY FOR NURSING CHAPTER 1Daniel Joshua MiguelPas encore d'évaluation
- Introduction to Microbes and Cellular BiologyDocument5 pagesIntroduction to Microbes and Cellular BiologyPauline PascuaDPas encore d'évaluation
- CellsDocument4 pagesCellsAIRENA RAIN TAMPOSPas encore d'évaluation
- Cell Structure and FunctionDocument32 pagesCell Structure and Functionnaveedalikhoso365Pas encore d'évaluation
- Human Cell AnatomyDocument56 pagesHuman Cell Anatomysanthiyasandy100% (2)
- Eukaryotic Cell StructureDocument24 pagesEukaryotic Cell StructureKarolina RyśkiewiczPas encore d'évaluation
- Cell Organelles & Their Functions 1112Document7 pagesCell Organelles & Their Functions 1112Amrul Zlalu AdaPas encore d'évaluation
- (Biochem) Chap 1 - Introduction To Biological CellsDocument35 pages(Biochem) Chap 1 - Introduction To Biological Cellslethithuhat67Pas encore d'évaluation
- 2 - CellDocument44 pages2 - CellNorie palmesPas encore d'évaluation
- III. Chapter 3 (Nucleus)Document5 pagesIII. Chapter 3 (Nucleus)rosalesjirehrPas encore d'évaluation
- Integrated Principles of ZoologyDocument41 pagesIntegrated Principles of ZoologyEstherPas encore d'évaluation
- Cell Organelles Structure & FunctionDocument4 pagesCell Organelles Structure & FunctionDaniel ChinPas encore d'évaluation
- (Organelles & Cellular Structures) H1 Bio NotesDocument11 pages(Organelles & Cellular Structures) H1 Bio Notesblah blehPas encore d'évaluation
- Cell Biology Inventory COMPLETEDocument18 pagesCell Biology Inventory COMPLETEmariam miladPas encore d'évaluation
- Reflective Journal For Module 2 Unit 1: Name: Yulo, Ires A. Section: BSED-SCI.3-BDocument6 pagesReflective Journal For Module 2 Unit 1: Name: Yulo, Ires A. Section: BSED-SCI.3-Bguirillita acutPas encore d'évaluation
- CellsDocument6 pagesCellsaryam aliPas encore d'évaluation
- CGEN - 100 - #2-Parts of A CellDocument2 pagesCGEN - 100 - #2-Parts of A CellEricka MasakayanPas encore d'évaluation
- CellsDocument25 pagesCellsAshwika RajanPas encore d'évaluation
- Organelles Chart FOR StudentDocument5 pagesOrganelles Chart FOR Studentnurul taqinah ismailPas encore d'évaluation
- NucleusDocument5 pagesNucleussiddharthr954Pas encore d'évaluation
- Molecular Biology and Diagnostic Intro To CytogeneticsDocument6 pagesMolecular Biology and Diagnostic Intro To Cytogeneticselijah montefalcoPas encore d'évaluation
- Microana Prelim ReviewerDocument17 pagesMicroana Prelim ReviewerjerlerositaPas encore d'évaluation
- Nucleus - Morphology and FunctionsDocument25 pagesNucleus - Morphology and FunctionsVytheeshwaran Vedagiri96% (24)
- Chapter 3. Cell Structure and Taxonomy ReviewerDocument7 pagesChapter 3. Cell Structure and Taxonomy ReviewerAlexandra MalagaPas encore d'évaluation
- 4, 5 - The Cytoplasm - 2Document58 pages4, 5 - The Cytoplasm - 2Muthana Bani YassenPas encore d'évaluation
- Cells The Living Units Part CDocument6 pagesCells The Living Units Part CDan Paulene AceboPas encore d'évaluation
- Reviewer in BiologyDocument16 pagesReviewer in BiologyAngel CastilloPas encore d'évaluation
- The Cell: Endomembrane System, Mitochondria, Chloroplasts, Cytoskeleton and Extracellular ComponentsDocument65 pagesThe Cell: Endomembrane System, Mitochondria, Chloroplasts, Cytoskeleton and Extracellular ComponentsMeljean Kalaw CastilloPas encore d'évaluation
- Cell Fucking MembraneDocument12 pagesCell Fucking MembraneColeen ParejaPas encore d'évaluation
- Burton's Microbiology For The Health Sciences: Chapter 3. Cell Structure and TaxonomyDocument25 pagesBurton's Microbiology For The Health Sciences: Chapter 3. Cell Structure and TaxonomyLindsay SwanepeolPas encore d'évaluation
- Burton's Microbiology For The Health Sciences: Chapter 3. Cell Structure and TaxonomyDocument25 pagesBurton's Microbiology For The Health Sciences: Chapter 3. Cell Structure and TaxonomyLindsay SwanepeolPas encore d'évaluation
- Cell Structure: A Guide to the OrganellesDocument11 pagesCell Structure: A Guide to the OrganellesAli MahmoudPas encore d'évaluation
- Science 6 Biology MidtermsDocument6 pagesScience 6 Biology MidtermsSamantha Avril UmandapPas encore d'évaluation
- Basic Building Blocks of Life Smallest Living Unit of An OrganismDocument2 pagesBasic Building Blocks of Life Smallest Living Unit of An OrganismkikomagsaysayPas encore d'évaluation
- CellDocument36 pagesCellwnslthmangulad05Pas encore d'évaluation
- BIO 345 S13 Continuity S14 (Chapter 5) S15 S16 S17 S18Document153 pagesBIO 345 S13 Continuity S14 (Chapter 5) S15 S16 S17 S18gaelle tannousPas encore d'évaluation
- BIO 266 - Cell Biology: Cells and OrganellesDocument78 pagesBIO 266 - Cell Biology: Cells and OrganellesVolodymyr KravchenkoPas encore d'évaluation
- MicroDocument5 pagesMicrotahashahzad9901Pas encore d'évaluation
- 3 - Cell Structures and Their FunctionsDocument12 pages3 - Cell Structures and Their FunctionsPrince TeodocioPas encore d'évaluation
- Lecture 3Document153 pagesLecture 3Miya GordPas encore d'évaluation
- 06-The Eukaryotic CellDocument76 pages06-The Eukaryotic Celldelia selmiPas encore d'évaluation
- Cell and Molecular BiologyDocument8 pagesCell and Molecular BiologySofia Christel JavierPas encore d'évaluation
- CellDocument27 pagesCellTom Anthony TonguiaPas encore d'évaluation
- Unit Mahluk Hidup - Penyusun Mahluk HidupDocument56 pagesUnit Mahluk Hidup - Penyusun Mahluk HidupYahya RhamadanPas encore d'évaluation
- Structure and Function of Cell: Presented by Miss - Robinsini.F 4 Year BSC Nursing Hosmat College of NursingDocument29 pagesStructure and Function of Cell: Presented by Miss - Robinsini.F 4 Year BSC Nursing Hosmat College of NursingJoshua SpotifyPas encore d'évaluation
- Cell Structure FunctionsDocument4 pagesCell Structure FunctionsGian Carlo MadriagaPas encore d'évaluation
- Cell Structure IVDocument119 pagesCell Structure IVDwi Puji AstiniPas encore d'évaluation
- Cell Structure IIIDocument27 pagesCell Structure IIIDwi Puji Astini100% (1)
- Cell Structure IDocument64 pagesCell Structure IDwi Puji Astini100% (1)
- Biologi - Scientific MethodsDocument6 pagesBiologi - Scientific MethodsDwi Puji AstiniPas encore d'évaluation
- Dwi Puji Astini - The Animalia Structure PresentationDocument75 pagesDwi Puji Astini - The Animalia Structure PresentationDwi Puji AstiniPas encore d'évaluation
- Biologi - Day3 Scientific MethodDocument13 pagesBiologi - Day3 Scientific MethodDwi Puji AstiniPas encore d'évaluation
- Biologi - Scientific MethodDocument13 pagesBiologi - Scientific MethodDwi Puji AstiniPas encore d'évaluation
- Cell Structure IIDocument35 pagesCell Structure IIDwi Puji AstiniPas encore d'évaluation
- Biologi - New Scientific MethodDocument21 pagesBiologi - New Scientific MethodDwi Puji AstiniPas encore d'évaluation
- Biologi - What Is LifeDocument8 pagesBiologi - What Is LifeDwi Puji AstiniPas encore d'évaluation
- Biologi - Biology Chapter 3Document19 pagesBiologi - Biology Chapter 3Dwi Puji Astini100% (1)
- Biologi - Biology Chapter 2Document26 pagesBiologi - Biology Chapter 2Dwi Puji AstiniPas encore d'évaluation
- Biologi - BiologyDocument6 pagesBiologi - BiologyDwi Puji AstiniPas encore d'évaluation
- Biologi - Biology Chapter 1Document12 pagesBiologi - Biology Chapter 1Dwi Puji Astini100% (3)
- Biologi - Biology Chapter 4Document17 pagesBiologi - Biology Chapter 4Dwi Puji Astini100% (1)
- Biologi - Biology Chapter 5Document18 pagesBiologi - Biology Chapter 5Dwi Puji AstiniPas encore d'évaluation
- Biologi - Biology Chapter 7Document19 pagesBiologi - Biology Chapter 7Dwi Puji Astini100% (2)
- Biologi - Biology Chapter 6Document17 pagesBiologi - Biology Chapter 6Dwi Puji AstiniPas encore d'évaluation
- Proceedings of The Xxi Annual Symposium of The Finnish Association of Mathematics and Science Education ResearchDocument611 pagesProceedings of The Xxi Annual Symposium of The Finnish Association of Mathematics and Science Education ResearchDwi Puji Astini100% (5)
- KA7630/KA7631: Fixed Multi-Output RegulatorDocument6 pagesKA7630/KA7631: Fixed Multi-Output RegulatorTecnico GenioPas encore d'évaluation
- Sports Project BbaDocument10 pagesSports Project Bbadonboy004Pas encore d'évaluation
- Today's Court Decision on Land Registration AppealDocument6 pagesToday's Court Decision on Land Registration AppealJohn Lester TanPas encore d'évaluation
- Is 2572 (2005) - Construction of Hollow and Solid Concrete Block Masonry - Code of PracticeDocument27 pagesIs 2572 (2005) - Construction of Hollow and Solid Concrete Block Masonry - Code of PracticeVIDYAPas encore d'évaluation
- Preparing Your Tracks For Mixing - SoundBetterDocument5 pagesPreparing Your Tracks For Mixing - SoundBetterEmmanuel TuffetPas encore d'évaluation
- Clay Minerals Including Related Phyllosilicates: Interdisciplinary Research and Inward IntegrationDocument16 pagesClay Minerals Including Related Phyllosilicates: Interdisciplinary Research and Inward Integrationbashir DarPas encore d'évaluation
- q2 w6 Asteroids Comets MeteorsDocument61 pagesq2 w6 Asteroids Comets MeteorsxenarealePas encore d'évaluation
- PractService Level Agreement NEBDN National Diploma in Dental NursingiceDocument21 pagesPractService Level Agreement NEBDN National Diploma in Dental NursingiceAdriana MihalachiPas encore d'évaluation
- Test BankDocument14 pagesTest BankB1111815167 WSBPas encore d'évaluation
- Pending 1667646640 ForeclosureSimulationDocument2 pagesPending 1667646640 ForeclosureSimulationpramod roy0% (1)
- Electromagnetic Spectrum 1 QP PDFDocument13 pagesElectromagnetic Spectrum 1 QP PDFWai HponePas encore d'évaluation
- Managing dextrose therapyDocument2 pagesManaging dextrose therapySanket TelangPas encore d'évaluation
- AirOS 3.4 - Ubiquiti Wiki#BasicWirelessSettings#BasicWirelessSettingsDocument24 pagesAirOS 3.4 - Ubiquiti Wiki#BasicWirelessSettings#BasicWirelessSettingsAusPas encore d'évaluation
- Different Types of AstrologyDocument16 pagesDifferent Types of AstrologyastrologerrajeevjiPas encore d'évaluation
- Section 4 QuizDocument2 pagesSection 4 QuizRizki Rian AnugrahaniPas encore d'évaluation
- Astrology and Athrishta K P 12 KS KrishnamurthiDocument401 pagesAstrology and Athrishta K P 12 KS Krishnamurthiaaditya80% (5)
- Elements of PoetryDocument50 pagesElements of PoetryBeing LoisPas encore d'évaluation
- Thomas McPherson Brown MD Treatment of Rheumatoid DiseaseDocument29 pagesThomas McPherson Brown MD Treatment of Rheumatoid DiseaseLidia Lidia100% (1)
- Legal CapacityDocument7 pagesLegal CapacityAreej UmerPas encore d'évaluation
- MLU Spec Sheet 250W 255WDocument2 pagesMLU Spec Sheet 250W 255WAdrian TurcanuPas encore d'évaluation
- First Masterclass Progress Test 1Document4 pagesFirst Masterclass Progress Test 1Billy Herington fullmasterPas encore d'évaluation
- English for Public Information LK 1Document7 pagesEnglish for Public Information LK 1jauhar muttaqinPas encore d'évaluation
- Electronics Engineering CDR SampleDocument7 pagesElectronics Engineering CDR SampleCDR Sample100% (1)
- MASAR - Volume 02Document214 pagesMASAR - Volume 02Khaled MamdouhPas encore d'évaluation
- T O E F L: Kaplan Analysis of Placement TestDocument1 pageT O E F L: Kaplan Analysis of Placement TestSharon DanielPas encore d'évaluation
- Simulink®Simulink Control Design User's GuideDocument1 352 pagesSimulink®Simulink Control Design User's GuideTe PekaPas encore d'évaluation
- The Metabolic and Endocrine Systems NotesDocument9 pagesThe Metabolic and Endocrine Systems NotesCarl Elexer Cuyugan Ano100% (2)
- Color Code Personality TestDocument4 pagesColor Code Personality TestJopsi100% (1)
- MT Level IDocument12 pagesMT Level Iidealparrot100% (1)
- 3 Strategic CapabilitiesDocument20 pages3 Strategic CapabilitiesMariyam HamdhaPas encore d'évaluation