Vous aimerez peut-être aussi
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (74)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- Review of Related LiteratureDocument6 pagesReview of Related LiteratureMaya Vil0% (1)
- Volpe's Understanding EvolutionDocument303 pagesVolpe's Understanding EvolutionAnonymous kg7YBMFH100% (1)
- The School of Allied Health Sciences ofDocument1 pageThe School of Allied Health Sciences ofMaya VilPas encore d'évaluation
- Basic Concepts of Computer Networks and Communications: Presented By: Joss Kendrick LiquidoDocument21 pagesBasic Concepts of Computer Networks and Communications: Presented By: Joss Kendrick LiquidoMaya VilPas encore d'évaluation
- Grading Sheet (Title Defense)Document1 pageGrading Sheet (Title Defense)Maya VilPas encore d'évaluation
- Cardiac RehabilitationDocument87 pagesCardiac RehabilitationMaya Vil100% (2)
- Sermon On The Ten Virgins - Parable of The Ten VirginsDocument4 pagesSermon On The Ten Virgins - Parable of The Ten VirginsMaya VilPas encore d'évaluation
- Postpartum Physical TherapyDocument10 pagesPostpartum Physical TherapyMaya VilPas encore d'évaluation
- Ankle and FootDocument188 pagesAnkle and FootMaya VilPas encore d'évaluation
- Presented By: Myra Leslie S. ViloriaDocument10 pagesPresented By: Myra Leslie S. ViloriaMaya VilPas encore d'évaluation
- Berg Balance Scale: By: Myra Leslie S. ViloriaDocument5 pagesBerg Balance Scale: By: Myra Leslie S. ViloriaMaya VilPas encore d'évaluation
- Clinical Case Study GonadsDocument18 pagesClinical Case Study GonadsMaya VilPas encore d'évaluation
- Bilateral OsteoarthritisDocument58 pagesBilateral OsteoarthritisMaya VilPas encore d'évaluation
- Adrenal Cortex Case StudyDocument21 pagesAdrenal Cortex Case StudyMaya Vil0% (1)
- LOW Back PainDocument111 pagesLOW Back PainMaya Vil100% (3)
- Congenital Malformation of The Brain: By: Myra Leslie S. Viloria References: Neuroimaging - Clinical ApplicationsDocument32 pagesCongenital Malformation of The Brain: By: Myra Leslie S. Viloria References: Neuroimaging - Clinical ApplicationsMaya VilPas encore d'évaluation
- Sensory AssessmentDocument3 pagesSensory AssessmentMaya VilPas encore d'évaluation
- Major Components of Social SciencesDocument27 pagesMajor Components of Social SciencesMaya VilPas encore d'évaluation
- Gender and Development Made EasyDocument49 pagesGender and Development Made EasyMaya Vil100% (1)
- Major Components of Social SciencesDocument18 pagesMajor Components of Social SciencesMaya VilPas encore d'évaluation
- Disorders of PigmentationDocument45 pagesDisorders of PigmentationMoayad NawaflehPas encore d'évaluation
- CELL CYCLE PresentationDocument22 pagesCELL CYCLE PresentationCalvin YusopPas encore d'évaluation
- What's in A Cell - Worksheet & ColoringDocument2 pagesWhat's in A Cell - Worksheet & ColoringNuria Tarancon0% (1)
- Diffusion and Osmosis Worksheet: Garrison O'Level Campus Cantt, QuettaDocument6 pagesDiffusion and Osmosis Worksheet: Garrison O'Level Campus Cantt, QuettagulminaPas encore d'évaluation
- Growth Kinetics 2014Document33 pagesGrowth Kinetics 2014Ivan Aulia BigbandPas encore d'évaluation
- CVS - Charts & DiagramsDocument20 pagesCVS - Charts & DiagramsMamathaPas encore d'évaluation
- Nursing ProcessDocument23 pagesNursing Processjulesubayubay5428100% (6)
- Hope Jennings - Dystopias of Matriarchal Power: Deconstructing The Womb in Angela Carter's Heroes and Villains and The Passion of New EveDocument11 pagesHope Jennings - Dystopias of Matriarchal Power: Deconstructing The Womb in Angela Carter's Heroes and Villains and The Passion of New Evedanutza123Pas encore d'évaluation
- Chapter4 Opt3Document30 pagesChapter4 Opt3Cza VerwinPas encore d'évaluation
- Instructions Template For Freshman ResumeDocument2 pagesInstructions Template For Freshman ResumeGuillaumeIPas encore d'évaluation
- Mob Ot 31753002739420Document276 pagesMob Ot 31753002739420fabriziozaraPas encore d'évaluation
- Eco Tourism DR Meeta Nihalani Kajol DhootDocument19 pagesEco Tourism DR Meeta Nihalani Kajol Dhootdr meetaPas encore d'évaluation
- 2011 Lecture 4b Ecosystem Dynamics IIDocument33 pages2011 Lecture 4b Ecosystem Dynamics IIJohn Zephyr TyskaPas encore d'évaluation
- Encyclopedia ES HealthDocument9 pagesEncyclopedia ES HealthGantsooj BPas encore d'évaluation
- Rolling Advertisement For FacultyDocument6 pagesRolling Advertisement For Facultymanahar bakshiPas encore d'évaluation
- Primo Vessels Dr. AshrafDocument7 pagesPrimo Vessels Dr. AshrafAshraful IslamPas encore d'évaluation
- Vitamin B6 Deficiency As A Cause of Polyneuropathy in POEMS Syndrome Rapid Recovery With Supplementation in Two CasesDocument7 pagesVitamin B6 Deficiency As A Cause of Polyneuropathy in POEMS Syndrome Rapid Recovery With Supplementation in Two CasesYaseen MohamnadPas encore d'évaluation
- Nanoparticles For Diagnostic Imaging andDocument12 pagesNanoparticles For Diagnostic Imaging andBrunaPas encore d'évaluation
- Biochemistry of GlycoproteinDocument6 pagesBiochemistry of GlycoproteinMahathir Mohmed100% (7)
- Botany B.SCDocument2 pagesBotany B.SCAkash Deep PandeyPas encore d'évaluation
- Daily Lesson Plan: 7e's ApproachDocument6 pagesDaily Lesson Plan: 7e's ApproachSonny MagdadaroPas encore d'évaluation
- Lecture 2 - ERTH 2403Document9 pagesLecture 2 - ERTH 2403Andre YimPas encore d'évaluation
- Clinical EMG ALSDocument7 pagesClinical EMG ALSAbul Shamsud DoulahPas encore d'évaluation
- BIO270 Prelab 1 Assignment 2014Document8 pagesBIO270 Prelab 1 Assignment 2014noahyuyanPas encore d'évaluation
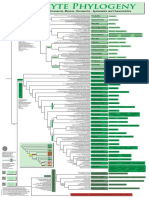
- Filogenia de Briofitos SensulatoDocument1 pageFilogenia de Briofitos SensulatoYuyitoS2714Pas encore d'évaluation
- (Brawer) Prostate Specific AntigenDocument342 pages(Brawer) Prostate Specific Antigenseeranga_raghuPas encore d'évaluation
- Ethylhexylglycerin Safety ToxicologyDocument17 pagesEthylhexylglycerin Safety Toxicologybfh83730Pas encore d'évaluation
- Being Left-Handed in A Right-Handed WorldDocument1 pageBeing Left-Handed in A Right-Handed Worldtranmee05Pas encore d'évaluation