Vous aimerez peut-être aussi
- Informe Desnaturalización Del Colageno - Gelatina de PataDocument5 pagesInforme Desnaturalización Del Colageno - Gelatina de PataDiego RangelPas encore d'évaluation
- Cambios Bioquimico PescadoDocument14 pagesCambios Bioquimico PescadoPaola Belen LlamocaPas encore d'évaluation
- Monografia de HuevosDocument10 pagesMonografia de HuevosHuarachi Rolin100% (1)
- Produccion de FrioDocument12 pagesProduccion de FriofrpatriciaPas encore d'évaluation
- Capitulo I ComposicionDocument14 pagesCapitulo I ComposicionSuarez Sulca Melanio100% (1)
- Solubilidad de ProteinasDocument8 pagesSolubilidad de ProteinasKaren Ocampo SerranoPas encore d'évaluation
- Principales Microorganismos Que Intervienen en La Fermentación AlcohólicaDocument4 pagesPrincipales Microorganismos Que Intervienen en La Fermentación AlcohólicaJoselynParedesPas encore d'évaluation
- Práctica 11 Leches Coaguladas - Enzimática y MixtaDocument7 pagesPráctica 11 Leches Coaguladas - Enzimática y MixtaJohan Camilo Restrepo ChandiPas encore d'évaluation
- Informe de GlutenDocument26 pagesInforme de GlutenOlmedo Calli OlveaPas encore d'évaluation
- INTRODUCCIÓNDocument9 pagesINTRODUCCIÓNCARLOS ALVA TICONAPas encore d'évaluation
- Fase Final Grupo1 4Document20 pagesFase Final Grupo1 4Alexa RMPas encore d'évaluation
- Informe BromatologiaDocument12 pagesInforme BromatologiaToño Vargas GutierrezPas encore d'évaluation
- PigmentosDocument11 pagesPigmentosRicky FasanandoPas encore d'évaluation
- Discusion de Vitamina CDocument1 pageDiscusion de Vitamina CGerzon Gonzales100% (1)
- PROBLEMAS DE BALANCE DE ENERGÍA 2011lDocument7 pagesPROBLEMAS DE BALANCE DE ENERGÍA 2011lalexander0% (1)
- Azúcar Convertida y CaramelizaciónDocument6 pagesAzúcar Convertida y CaramelizaciónJoao JachoPas encore d'évaluation
- Difusividad en GranosDocument10 pagesDifusividad en GranosYoryi FloresPas encore d'évaluation
- Determinacion de Extracto EtereoDocument5 pagesDeterminacion de Extracto EtereoEdwin A. Aguilar M.Pas encore d'évaluation
- Resultados y DiscusionesDocument3 pagesResultados y DiscusionesDeysi MelisaPas encore d'évaluation
- Lab. 12 CARAMELIZACION EN MEDIOS ACIDOS Y ALCALINOSDocument2 pagesLab. 12 CARAMELIZACION EN MEDIOS ACIDOS Y ALCALINOSFanny C. Diaz DelgadoPas encore d'évaluation
- Pardeamiento No Enzimático en Productos AgroindustrialesDocument17 pagesPardeamiento No Enzimático en Productos AgroindustrialesAlexander Yauri GomezPas encore d'évaluation
- Práctica #01Document14 pagesPráctica #01Alexander Yauri GomezPas encore d'évaluation
- Importancia de La LactosaDocument11 pagesImportancia de La LactosaMarioly Rodriguez VidalPas encore d'évaluation
- Agua en Los Alimento Liquidos, Solidos, y ColoidesDocument13 pagesAgua en Los Alimento Liquidos, Solidos, y ColoidesginoPas encore d'évaluation
- Analisis Del HuevoDocument9 pagesAnalisis Del HuevoMónica Del Rio ZuñigaPas encore d'évaluation
- Contaminación, Conservación y Ateración de La Leche y DerivadosDocument7 pagesContaminación, Conservación y Ateración de La Leche y Derivadoskelly rosarioPas encore d'évaluation
- Analisis de Pan IntroduccionDocument7 pagesAnalisis de Pan IntroduccionJade Anny QSPas encore d'évaluation
- Informe Leche ProteinaDocument3 pagesInforme Leche ProteinaElmerLeninSanchezRamosPas encore d'évaluation
- DiscusionesDocument4 pagesDiscusionesTatiana YanquiPas encore d'évaluation
- Computo AminoacidicoDocument4 pagesComputo AminoacidicoKaren LemaPas encore d'évaluation
- Determinacion Humedad AlimentosDocument17 pagesDeterminacion Humedad AlimentosAnthony AyalaPas encore d'évaluation
- Segundo Informe de BioquimicaDocument3 pagesSegundo Informe de Bioquimicamaximo palomino rodasPas encore d'évaluation
- Expo Bioquimica de Hortalizas ImprimirDocument57 pagesExpo Bioquimica de Hortalizas ImprimirYoel Perez Herrera100% (1)
- Charqui Cordero Al 25 %Document7 pagesCharqui Cordero Al 25 %Milita BCPas encore d'évaluation
- INFORME de El HeladoDocument17 pagesINFORME de El HeladoJonathan RobayoPas encore d'évaluation
- Harina de PlatanoDocument8 pagesHarina de PlatanoVanessa Paulino GarciaPas encore d'évaluation
- Propiedades Del YogurtDocument34 pagesPropiedades Del YogurtirisPas encore d'évaluation
- Pardeamiento Enzimatica y No Enzimatico en Los AlimentosDocument21 pagesPardeamiento Enzimatica y No Enzimatico en Los AlimentosThalia Vasquez Vega100% (1)
- Informe Control de Calidad Laboratorio 3Document22 pagesInforme Control de Calidad Laboratorio 3Pablo Andrés Morán Santibáñez0% (1)
- Informe Alimentos de IV GamaDocument11 pagesInforme Alimentos de IV GamaKlaudia VieraPas encore d'évaluation
- Tarea 04Document6 pagesTarea 04AVALOS WONG MILAGROS ISABELPas encore d'évaluation
- Informe SalchichaDocument31 pagesInforme SalchichaWagner Gamboa CruzadoPas encore d'évaluation
- Determinacion de Humedad de Los AlimentosDocument9 pagesDeterminacion de Humedad de Los AlimentosRichard Klever Santos AlarconPas encore d'évaluation
- Introduccion PetipanDocument4 pagesIntroduccion PetipanBenjamin Umiyauri100% (1)
- Tarea - Práctica 04 - Análisis de Carnes - PH, Acidez y AdulteracionesDocument6 pagesTarea - Práctica 04 - Análisis de Carnes - PH, Acidez y AdulteracionesCoraima GuevaraPas encore d'évaluation
- CASEINADocument17 pagesCASEINAWalt Tu Amixito QeridPas encore d'évaluation
- Tarea 01Document6 pagesTarea 01AVALOS WONG MILAGROS ISABEL100% (2)
- CalorimetríaDocument7 pagesCalorimetríaCinthya NorabuenaPas encore d'évaluation
- Informe 03 PanificacionDocument10 pagesInforme 03 PanificacionMit TorresPas encore d'évaluation
- Capituo Xi Acidez Total Titulable PDFDocument26 pagesCapituo Xi Acidez Total Titulable PDFMaria Guadalupe Yaranga AvendañoPas encore d'évaluation
- Determinación de Lactosa - Grupo 1Document15 pagesDeterminación de Lactosa - Grupo 1rolandoPas encore d'évaluation
- Marco Teorico-Practica 1Document5 pagesMarco Teorico-Practica 1Luzmila CamaPas encore d'évaluation
- Control de Calidad de La LecheDocument9 pagesControl de Calidad de La Lechejosseline adriana rivera revollo100% (1)
- Teoria de MorcillaDocument2 pagesTeoria de MorcillaLalo la LocuraPas encore d'évaluation
- Analisis BromatologicoDocument7 pagesAnalisis Bromatologicoerick callenova mPas encore d'évaluation
- Informe Listo de YogurtDocument16 pagesInforme Listo de YogurtomarPas encore d'évaluation
- Resumen Fisiologia Aplicada A La Educacion Fisica - CelulaDocument26 pagesResumen Fisiologia Aplicada A La Educacion Fisica - CelulaManuel HermidaPas encore d'évaluation
- Clase 5Document11 pagesClase 5Chávez Meza RodrigoPas encore d'évaluation
- Energética de Contraccion MuscularDocument7 pagesEnergética de Contraccion MuscularFlorr VerasayPas encore d'évaluation
- Transmision NeuromuscularDocument23 pagesTransmision Neuromuscularedward_10Pas encore d'évaluation
- La EncuestaDocument11 pagesLa EncuestaAngela Valencia ChavezPas encore d'évaluation
- Violencia SexualDocument23 pagesViolencia SexualAngela Valencia ChavezPas encore d'évaluation
- Principales AccionesDocument3 pagesPrincipales AccionesAngela Valencia ChavezPas encore d'évaluation
- Trabajos Modelos Calidad Salud MexicoDocument5 pagesTrabajos Modelos Calidad Salud MexicoAngela Valencia ChavezPas encore d'évaluation
- Angela Valencia Chavez Poder de Los GerentesDocument8 pagesAngela Valencia Chavez Poder de Los GerentesAngela Valencia ChavezPas encore d'évaluation
- Vigilancia de La Calidad de Agua para ConsumoDocument10 pagesVigilancia de La Calidad de Agua para ConsumoAngela Valencia ChavezPas encore d'évaluation
- Silicon ValleyDocument3 pagesSilicon ValleyAngela Valencia ChavezPas encore d'évaluation
- CALAMARDocument6 pagesCALAMARAngela Valencia ChavezPas encore d'évaluation
- Paladar DuroDocument3 pagesPaladar DuroAngela Valencia ChavezPas encore d'évaluation
- Ecologia PoliticaDocument13 pagesEcologia PoliticaAngela Valencia ChavezPas encore d'évaluation
- Decision EficazDocument7 pagesDecision EficazAngela Valencia ChavezPas encore d'évaluation
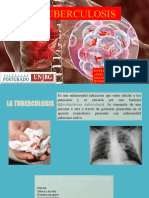
- 'Exposicion TBCDocument12 pages'Exposicion TBCAngela Valencia ChavezPas encore d'évaluation
- Ensayo EcofilosofiaDocument12 pagesEnsayo EcofilosofiaAngela Valencia ChavezPas encore d'évaluation
- CALAMARDocument6 pagesCALAMARAngela Valencia ChavezPas encore d'évaluation
- Historia de La Enfermedad TUBERCULOSISDocument4 pagesHistoria de La Enfermedad TUBERCULOSISAngela Valencia ChavezPas encore d'évaluation
- Trabajo de Aleaciones MetalicasDocument6 pagesTrabajo de Aleaciones MetalicasAngela Valencia ChavezPas encore d'évaluation
- Tuberculosis EnteroperitonealDocument18 pagesTuberculosis EnteroperitonealAngela Valencia Chavez0% (1)
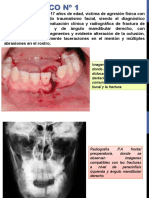
- Fracturas MandibulresDocument14 pagesFracturas MandibulresAngela Valencia ChavezPas encore d'évaluation
- HerniasDocument13 pagesHerniasAngela Valencia ChavezPas encore d'évaluation
- AlcoholismoDocument20 pagesAlcoholismoAngela Valencia ChavezPas encore d'évaluation
- Salud Oral, Lavado de ManosDocument7 pagesSalud Oral, Lavado de ManosAngela Valencia ChavezPas encore d'évaluation
- Tecnología Pesquera BásicaDocument15 pagesTecnología Pesquera BásicaAngela Valencia ChavezPas encore d'évaluation
- QueilosDocument4 pagesQueilosAngela Valencia Chavez100% (1)
- INTRAOPERATORIO Molares SuperioresDocument12 pagesINTRAOPERATORIO Molares SuperioresAngela Valencia ChavezPas encore d'évaluation
- Patrimonio CulturalDocument13 pagesPatrimonio CulturalAngela Valencia ChavezPas encore d'évaluation
- Resumen Aislamiento AbsolutoDocument3 pagesResumen Aislamiento AbsolutoAngela Valencia Chavez100% (1)
- Homosexual I DadDocument23 pagesHomosexual I DadAngela Valencia ChavezPas encore d'évaluation
- Tecnica Aaplicación en Conjunto La Goma de Dique Con El Clamp y Después El ArcoDocument3 pagesTecnica Aaplicación en Conjunto La Goma de Dique Con El Clamp y Después El ArcoAngela Valencia ChavezPas encore d'évaluation
- Analisis de Tejidos Blandos AyalaDocument3 pagesAnalisis de Tejidos Blandos AyalaAngela Valencia ChavezPas encore d'évaluation
- Acidos Grasos AnchovetaDocument2 pagesAcidos Grasos AnchovetaAngela Valencia ChavezPas encore d'évaluation
- Difusividad Marco TeoricoDocument13 pagesDifusividad Marco TeoricoJavixiitoOPas encore d'évaluation
- Semana 10 - PAE en Trastornos o Complicaciones de OtorrinolaringologiaDocument4 pagesSemana 10 - PAE en Trastornos o Complicaciones de OtorrinolaringologiaAriana LizbethPas encore d'évaluation
- BiologiaDocument21 pagesBiologiaBadr AbdounPas encore d'évaluation
- Tesis de Grado Radiologia e ImagenologíaDocument41 pagesTesis de Grado Radiologia e Imagenologíakleiverdd12Pas encore d'évaluation
- Hds AcetilenoDocument6 pagesHds Acetilenonancy toro toroPas encore d'évaluation
- Ud3 AbafsmDocument30 pagesUd3 AbafsmAntonio Rocio Pradas E IbanezPas encore d'évaluation
- Tabla PeriodicaDocument14 pagesTabla PeriodicaDanielPas encore d'évaluation
- La Publicidad ActividadesDocument4 pagesLa Publicidad ActividadesKarla OrtizPas encore d'évaluation
- Cuestionario de Productos FermentadosDocument6 pagesCuestionario de Productos Fermentadosjosseline adriana rivera revolloPas encore d'évaluation
- Taller 2. Método Algebraico, Simplex, Dos Fases y Gran M - COMPLETO PDFDocument9 pagesTaller 2. Método Algebraico, Simplex, Dos Fases y Gran M - COMPLETO PDFJavier VilladaPas encore d'évaluation
- SILABO Operatividad Maquina PDFDocument7 pagesSILABO Operatividad Maquina PDFCesarPercyNavarroPas encore d'évaluation
- Epidemiologia EspacialDocument20 pagesEpidemiologia Espacialbacanisimo10Pas encore d'évaluation
- Actividad Sobre DolorDocument9 pagesActividad Sobre DolorRomina SterponePas encore d'évaluation
- Regeneración Ósea Guiada Simultánea A La Colocación de Implantes: Presentación de Un Caso ClínicoDocument8 pagesRegeneración Ósea Guiada Simultánea A La Colocación de Implantes: Presentación de Un Caso ClínicoSABUESO FINANCIEROPas encore d'évaluation
- Siga (Sistema Integrado de Gestion AdministrativaDocument13 pagesSiga (Sistema Integrado de Gestion AdministrativaFrachescoly Lima QuispePas encore d'évaluation
- Texto GeneradoDocument1 pageTexto GeneradoLimbic LimbicPas encore d'évaluation
- Técnicas OperativasDocument2 pagesTécnicas OperativaswoonsmartinPas encore d'évaluation
- LIBERACON DE FARMACOS y ALIMENTOS EN ABSORCION DE FARMACOSDocument109 pagesLIBERACON DE FARMACOS y ALIMENTOS EN ABSORCION DE FARMACOSRoxana Magaly Chavez PillacaPas encore d'évaluation
- Chapter 7 - Enzymatic Quorum Quenching in Biofilms - 2019 - Quorum SensingDocument21 pagesChapter 7 - Enzymatic Quorum Quenching in Biofilms - 2019 - Quorum SensingFrancelia Quiñonez RuvalcabaPas encore d'évaluation
- Capacitacion PersonalDocument26 pagesCapacitacion PersonalDouglas UribePas encore d'évaluation
- 370LDocument695 pages370LPaul SmithPas encore d'évaluation
- Triptico Reciclar Papel - Ariana Diaz C.Document2 pagesTriptico Reciclar Papel - Ariana Diaz C.Ariana Francesca Diaz CabrejosPas encore d'évaluation
- Cuadro Filosofía Esa Búsqueda ReflexivaDocument7 pagesCuadro Filosofía Esa Búsqueda ReflexivaJoaquin RiverosPas encore d'évaluation
- Studio One 3 Guía de Inicio Rápido PDFDocument19 pagesStudio One 3 Guía de Inicio Rápido PDFLuiz Henrique MendesPas encore d'évaluation
- Presentacion Final Fundamentos DinamoDocument36 pagesPresentacion Final Fundamentos DinamoAdriánPechCerveraPas encore d'évaluation
- Mejoras PackingDocument2 pagesMejoras PackingAngela Fuentes higueraPas encore d'évaluation
- RS PCD 0842022 PDFDocument3 pagesRS PCD 0842022 PDFJesus Leonidas Callata OliveraPas encore d'évaluation
- El Exceso en Las Causas de JustificacionDocument108 pagesEl Exceso en Las Causas de JustificacionErik Manuel Vasquez LlerenaPas encore d'évaluation
- Anamnesis Preclinica PDFDocument3 pagesAnamnesis Preclinica PDFDANITZA LIDIANETH CARRILLO HERRERAPas encore d'évaluation
- Manual de CiclismoDocument19 pagesManual de CiclismoJuegos RosarioPas encore d'évaluation