Vous aimerez peut-être aussi
- Basic EKG ECG Rhythms CheatsheetDocument1 pageBasic EKG ECG Rhythms CheatsheetAhmad83% (6)
- EKG/ECG Interpretation Made Easy: A Practical Approach to Passing the ECG/EKG Portion of NCLEXD'EverandEKG/ECG Interpretation Made Easy: A Practical Approach to Passing the ECG/EKG Portion of NCLEXÉvaluation : 5 sur 5 étoiles5/5 (2)
- Review Notes 2000 - CardiologyDocument58 pagesReview Notes 2000 - Cardiologyeliaszavaleta100% (1)
- Ekg Full BibleDocument6 pagesEkg Full BibleTJPas encore d'évaluation
- Signals & Systems - NOTESDocument56 pagesSignals & Systems - NOTESTaimur Musharraf91% (70)
- Learning ECGDocument187 pagesLearning ECGPhysiology by Dr Raghuveer100% (2)
- Risk Factors & Prognosis For USMLE STEP 3 PDFDocument182 pagesRisk Factors & Prognosis For USMLE STEP 3 PDFSadia GhaniPas encore d'évaluation
- Blood Circulation System 10 PDFDocument29 pagesBlood Circulation System 10 PDFSardar jiPas encore d'évaluation
- Clinical Guidelines in Primary Care 3rdDocument12 pagesClinical Guidelines in Primary Care 3rdmimi100% (1)
- BiopotentialsDocument30 pagesBiopotentialsFidaa Jaafrah100% (2)
- Cardiac Considerations in Chronic Lung Disease 2020Document276 pagesCardiac Considerations in Chronic Lung Disease 2020Dani CapiPas encore d'évaluation
- PASS - Coursebook MAINDocument529 pagesPASS - Coursebook MAINkabal0850% (2)
- Nursing Test 2 (NP Iii)Document19 pagesNursing Test 2 (NP Iii)Yuxin LiuPas encore d'évaluation
- ECG Interpretation in Small AnimalsDocument10 pagesECG Interpretation in Small Animalsgacf1974Pas encore d'évaluation
- BIORESONANCE EnglishDocument12 pagesBIORESONANCE EnglishJose La100% (1)
- Azin Alizadehasl, Majid Maleki - Case-Based Clinical Cardiology-Springer (2021)Document480 pagesAzin Alizadehasl, Majid Maleki - Case-Based Clinical Cardiology-Springer (2021)Phạm Anh Duy100% (1)
- Toroidal Structure of The Assemblage Point AppendixDocument5 pagesToroidal Structure of The Assemblage Point AppendixminteiluminataPas encore d'évaluation
- Arrythmia Name: Normal Sinus RhythmDocument7 pagesArrythmia Name: Normal Sinus RhythmJulx0Pas encore d'évaluation
- What Is Qi Energy Flow Interpretation of Our Results of Ami Measurements Naohiro Nagayama (Subtle Energies, Vol 21, No 3)Document10 pagesWhat Is Qi Energy Flow Interpretation of Our Results of Ami Measurements Naohiro Nagayama (Subtle Energies, Vol 21, No 3)Cambiador de MundoPas encore d'évaluation
- Ventricular ArrhytmiaDocument30 pagesVentricular ArrhytmiaIkhsan AmadeaPas encore d'évaluation
- Clinical Demonstration: Topic: Electrocardiogram (ECG)Document24 pagesClinical Demonstration: Topic: Electrocardiogram (ECG)soniya josephPas encore d'évaluation
- Taibah University Interns: Umm Al - Qura University SLE Questions 2 EditionDocument531 pagesTaibah University Interns: Umm Al - Qura University SLE Questions 2 EditionFammo Moidu100% (2)
- Eggermont 2019 ABRDocument15 pagesEggermont 2019 ABRSujeet PathakPas encore d'évaluation
- OF THE York Academy: Study Activity SingleDocument18 pagesOF THE York Academy: Study Activity Singleedycs10Pas encore d'évaluation
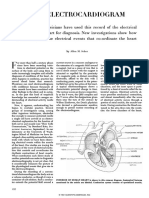
- The Electrocardiogram: Allen M. ScherDocument12 pagesThe Electrocardiogram: Allen M. ScherrkukgPas encore d'évaluation
- 38.17 Cardiac Pacing: Basic Concepts: Renato Pietro RicciDocument16 pages38.17 Cardiac Pacing: Basic Concepts: Renato Pietro RicciurtikikePas encore d'évaluation
- 202 Full-Text PDFDocument7 pages202 Full-Text PDFlionkingPas encore d'évaluation
- John Eccles SynapsesDocument22 pagesJohn Eccles SynapsesLeandro XimonPas encore d'évaluation
- CH 5 ElectricityDocument39 pagesCH 5 ElectricityAbdullahi Mohamed IsakPas encore d'évaluation
- BM Unit-IIIDocument12 pagesBM Unit-IIIHp KPas encore d'évaluation
- Learning Objectives: Chapter 4: Electrophysiology of The HeartDocument28 pagesLearning Objectives: Chapter 4: Electrophysiology of The HeartLaura PuchePas encore d'évaluation
- Ecgs Made Easy 6th Edition Aehlert Test BankDocument16 pagesEcgs Made Easy 6th Edition Aehlert Test Bankpatriciaoneillozmpwqbdra100% (14)
- Grupo 1 Practica 14Document12 pagesGrupo 1 Practica 14Sadit Chauca VelaPas encore d'évaluation
- A Fib 3Document9 pagesA Fib 3palak32Pas encore d'évaluation
- Bioinstrumentación - Pres3Document43 pagesBioinstrumentación - Pres3daniela0422Pas encore d'évaluation
- 1985 DendriticspinesDocument4 pages1985 Dendriticspinesangelamm1Pas encore d'évaluation
- Effect of Current Flow - 1951Document10 pagesEffect of Current Flow - 1951Владимир ДружининPas encore d'évaluation
- Cardiac Electrophysiology Using Ls Dyna R PDFDocument15 pagesCardiac Electrophysiology Using Ls Dyna R PDFЮрий НовожиловPas encore d'évaluation
- The Australian Journal of Physiotherapy: Electrodiagnostic TechniquesDocument4 pagesThe Australian Journal of Physiotherapy: Electrodiagnostic TechniquesumangPas encore d'évaluation
- Neuropatias Periféricas - 2007Document312 pagesNeuropatias Periféricas - 2007Claudio LopesPas encore d'évaluation
- 1982 Synapses Between L-Neurons J Comp PhysiolDocument14 pages1982 Synapses Between L-Neurons J Comp PhysiolClub OrilyonPas encore d'évaluation
- Heart PhysiolDocument84 pagesHeart PhysiolChintya Ayu ChampakaPas encore d'évaluation
- Electrical Resonance With Voltage-Gated Ion ChannelsDocument8 pagesElectrical Resonance With Voltage-Gated Ion ChannelsNusrat JahanPas encore d'évaluation
- 05 Biomedical EngineeringDocument357 pages05 Biomedical EngineeringAngel Celestial100% (1)
- Brain ResearchDocument9 pagesBrain ResearchKarina SjPas encore d'évaluation
- Neurophysiologic Basis of EEG: Piotr OlejniczakDocument4 pagesNeurophysiologic Basis of EEG: Piotr OlejniczakioanviPas encore d'évaluation
- BiosignalDocument27 pagesBiosignalAurongo NasirPas encore d'évaluation
- CH 4 FALL 15-16 Section 3-10Document75 pagesCH 4 FALL 15-16 Section 3-10giridharsinghPas encore d'évaluation
- The Magnetocardiogram A New Approach To The Fields Surrounding The HeartDocument6 pagesThe Magnetocardiogram A New Approach To The Fields Surrounding The Heartkpgs12Pas encore d'évaluation
- ECG Lectures 2023-2024Document29 pagesECG Lectures 2023-2024owronrawan74Pas encore d'évaluation
- Geddes 2004Document6 pagesGeddes 2004Kevin SzePas encore d'évaluation
- Dwnload Full Ecgs Made Easy 6th Edition Aehlert Test Bank PDFDocument35 pagesDwnload Full Ecgs Made Easy 6th Edition Aehlert Test Bank PDFabetterguerillal4o2100% (14)
- Clinical Electrocardiography and ECG Interpretation - ECG & ECHODocument15 pagesClinical Electrocardiography and ECG Interpretation - ECG & ECHORimaPas encore d'évaluation
- ECG CardiotachometerDocument43 pagesECG CardiotachometerWillius David DjoPas encore d'évaluation
- Roth-MuscleNerve 1990Document9 pagesRoth-MuscleNerve 1990Simran ChoudharyPas encore d'évaluation
- Blume 2004Document8 pagesBlume 2004jela1309Pas encore d'évaluation
- 2 Marks Q & ADocument11 pages2 Marks Q & AJayaprakash JayaramanPas encore d'évaluation
- Electrical Activity of Excitable Cells: The Resting StateDocument19 pagesElectrical Activity of Excitable Cells: The Resting State정은규Pas encore d'évaluation
- April 2008Document11 pagesApril 2008lvrevathiPas encore d'évaluation
- Cellule EcetabileDocument8 pagesCellule EcetabileAriel OlshevskyPas encore d'évaluation
- 247 FullDocument6 pages247 FullBryan FarrelPas encore d'évaluation
- History of VectocardiographyDocument31 pagesHistory of VectocardiographyMarcos JacomePas encore d'évaluation
- EI2311-Biomedical Instrumentation Question BankDocument20 pagesEI2311-Biomedical Instrumentation Question BankrajapandiyaPas encore d'évaluation
- AmericaDocument4 pagesAmericaJOSE HERNANDO JOYA HERERAPas encore d'évaluation
- BMI 2marksDocument8 pagesBMI 2marksPradheep RajanPas encore d'évaluation
- Ch-9 Electricity Within The BodyDocument55 pagesCh-9 Electricity Within The BodyLindsey AdalsonPas encore d'évaluation
- Detection of Endogenous Electromagnetic FieldDocument9 pagesDetection of Endogenous Electromagnetic FieldAlexandruPas encore d'évaluation
- Ecgs Made Easy 6th Edition Aehlert Test BankDocument35 pagesEcgs Made Easy 6th Edition Aehlert Test Bankconstanceholmesz7jjy100% (17)
- KIN 270 Unit 2 Neural Physiology Part 1Document4 pagesKIN 270 Unit 2 Neural Physiology Part 1Armando AlehandroPas encore d'évaluation
- Powerlab: Physiology of The Nerve: KeywordsDocument7 pagesPowerlab: Physiology of The Nerve: KeywordsCJ RanesesPas encore d'évaluation
- Evidence For A Primitive DC Electrical Analog System Controlling Brain FunctionDocument18 pagesEvidence For A Primitive DC Electrical Analog System Controlling Brain FunctionordoPas encore d'évaluation
- Electromyogram - Principles and ApplicationsDocument12 pagesElectromyogram - Principles and ApplicationsIoana PetrePas encore d'évaluation
- Endocardial Infarct Scar Recognition by MyocardialDocument8 pagesEndocardial Infarct Scar Recognition by MyocardialMihai-Marian RuseiPas encore d'évaluation
- ECG Feature Extraction For Stress Recognition in Automobile DriversDocument5 pagesECG Feature Extraction For Stress Recognition in Automobile DriversVeronica JanethPas encore d'évaluation
- EI6704 2marksDocument12 pagesEI6704 2marksssignnPas encore d'évaluation
- On The Measurement of Electrical Impedance Spectroscopy (EIS) of The Human HeadDocument15 pagesOn The Measurement of Electrical Impedance Spectroscopy (EIS) of The Human HeadgpaivPas encore d'évaluation
- Neurobiology of Motor Control: Fundamental Concepts and New DirectionsD'EverandNeurobiology of Motor Control: Fundamental Concepts and New DirectionsScott L. HooperPas encore d'évaluation
- Brain-Computer Interfaces 1: Methods and PerspectivesD'EverandBrain-Computer Interfaces 1: Methods and PerspectivesMaureen ClercPas encore d'évaluation
- 00855497Document7 pages00855497taharnebePas encore d'évaluation
- Novel DCS Transmitter and Receiver AntennasDocument29 pagesNovel DCS Transmitter and Receiver AntennasVijayRajPas encore d'évaluation
- Planar L-Strip Fed Broadband Micro Strip AntennaDocument3 pagesPlanar L-Strip Fed Broadband Micro Strip AntennaPratik MevadaPas encore d'évaluation
- Tamil Nadu Service RulesDocument100 pagesTamil Nadu Service RulesJoswa Santhakumar100% (1)
- Impedance Transformation and Impedance MatchingDocument36 pagesImpedance Transformation and Impedance MatchingThivaPas encore d'évaluation
- A Wideband Rectangular Microstrip Antenna Using An Asymmetric T-Shaped FeedDocument4 pagesA Wideband Rectangular Microstrip Antenna Using An Asymmetric T-Shaped FeedVijayRajPas encore d'évaluation
- Section 5 1 Matching With Lumped Elements PackageDocument29 pagesSection 5 1 Matching With Lumped Elements PackageVijayRajPas encore d'évaluation
- Seating Arrangement Part01: Problem: 01Document2 pagesSeating Arrangement Part01: Problem: 01VijayRajPas encore d'évaluation
- Caspers S ParametersDocument54 pagesCaspers S ParametersVijayRajPas encore d'évaluation
- Medical ElectronicsDocument3 pagesMedical ElectronicsVijayRajPas encore d'évaluation
- Fiber ConnectorsDocument29 pagesFiber ConnectorsVijayRaj100% (1)
- Answer: (C) : Q. No. 1 - 5 Carry One Mark EachDocument30 pagesAnswer: (C) : Q. No. 1 - 5 Carry One Mark EachSunny Dara Rinnah SusanthPas encore d'évaluation
- Fiber SplicesDocument19 pagesFiber SplicesVijayRajPas encore d'évaluation
- RF Circuit Design - Theory and ApplicationsDocument653 pagesRF Circuit Design - Theory and ApplicationsAyman Barakat94% (18)
- Seating Arrangement Part02: Problem: 01Document2 pagesSeating Arrangement Part02: Problem: 01VijayRajPas encore d'évaluation
- A Project Report at Cell Phone DetectorDocument83 pagesA Project Report at Cell Phone DetectorDrishti JainPas encore d'évaluation
- RF May June 2015 QPDocument2 pagesRF May June 2015 QPVijayRajPas encore d'évaluation
- History of SDocument4 pagesHistory of SVijayRajPas encore d'évaluation
- Coding and Decoding Questions Shortcut Tricks in Reasoning of AptitudeDocument18 pagesCoding and Decoding Questions Shortcut Tricks in Reasoning of AptitudeVijayRajPas encore d'évaluation
- EE407 Lec 1 2 1Document13 pagesEE407 Lec 1 2 1VijayRajPas encore d'évaluation
- ch16 TwoportnetworksDocument20 pagesch16 TwoportnetworksVijayRajPas encore d'évaluation
- JUN13Document3 pagesJUN13Anonymous lt2LFZHPas encore d'évaluation
- Microwave Engineering Ec 432Document25 pagesMicrowave Engineering Ec 432ainugiriPas encore d'évaluation
- RF and Micro Nov Dec 12Document2 pagesRF and Micro Nov Dec 12VijayRajPas encore d'évaluation
- EC2403 Ec2403 RF and Microwave EnggDocument7 pagesEC2403 Ec2403 RF and Microwave EnggVijayRajPas encore d'évaluation
- RF and Micro Nov Dec 12Document2 pagesRF and Micro Nov Dec 12VijayRajPas encore d'évaluation
- RF May June 2015 QPDocument2 pagesRF May June 2015 QPVijayRajPas encore d'évaluation
- When To Treat Extrasystole - Dr. RerdinDocument51 pagesWhen To Treat Extrasystole - Dr. RerdinStanley GunawanPas encore d'évaluation
- Care of Clients With Physiologic and Psychosocial Alterations - Doc Version 1Document46 pagesCare of Clients With Physiologic and Psychosocial Alterations - Doc Version 1CLAIRE VillaviejaPas encore d'évaluation
- Patofisiologi Sistem KardiovaskularDocument157 pagesPatofisiologi Sistem KardiovaskularNyanmaruPas encore d'évaluation
- ECG Arrhythmia Classification Using Deep Convolution Neural Networks in Transfer LearningDocument54 pagesECG Arrhythmia Classification Using Deep Convolution Neural Networks in Transfer LearningDayana VieiraPas encore d'évaluation
- OpenShift - Container - Platform 4.3 Storage en USDocument80 pagesOpenShift - Container - Platform 4.3 Storage en USOktavian AugustusPas encore d'évaluation
- ECG Physician's Guide (Japanese)Document49 pagesECG Physician's Guide (Japanese)Mohammad NasriPas encore d'évaluation
- Baby Bear Book 3rd Edition - JAN2016Document344 pagesBaby Bear Book 3rd Edition - JAN2016lim_kai_weiPas encore d'évaluation
- Ecg For AnkiDocument3 pagesEcg For AnkijPas encore d'évaluation
- EKG Interpretasi Dan Lethal Aritmia: Aan NuraeniDocument34 pagesEKG Interpretasi Dan Lethal Aritmia: Aan NuraeniCitra Marchelina Novilini100% (1)
- QuestionsDocument26 pagesQuestionsLyka Mae Imbat - PacnisPas encore d'évaluation
- Gabbe's Obstetrics Chapter 3 gm5.0Document30 pagesGabbe's Obstetrics Chapter 3 gm5.0AnniePas encore d'évaluation
- 71Document27 pages71onuchukwu chibuzorPas encore d'évaluation
- Dental Management of The Patient With Cardiac Arrhythmias: An UpdateDocument5 pagesDental Management of The Patient With Cardiac Arrhythmias: An UpdateDiego Zúñiga de LeónPas encore d'évaluation
- Management of Patients With Dysrhythmias and Conduction Problems DysrhythmiasDocument15 pagesManagement of Patients With Dysrhythmias and Conduction Problems DysrhythmiasRoyce Vincent TizonPas encore d'évaluation