Vous aimerez peut-être aussi
- TP Microscopios-2011Document3 pagesTP Microscopios-2011Bastián MacKensyPas encore d'évaluation
- Práctica Docente IDocument6 pagesPráctica Docente IMariaLujanBustosPas encore d'évaluation
- PDFDocument2 pagesPDFMariaLujanBustosPas encore d'évaluation
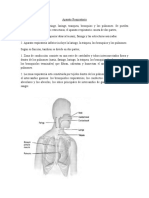
- Aparato RespiratorioDocument18 pagesAparato RespiratorioMariaLujanBustosPas encore d'évaluation
- Envoltura Nuclear y Complejo Del Poro NuDocument24 pagesEnvoltura Nuclear y Complejo Del Poro NuMariaLujanBustosPas encore d'évaluation
- Informe Zoocriaderos de Avestruces y MadDocument19 pagesInforme Zoocriaderos de Avestruces y Madyovanny antonPas encore d'évaluation
- Naturales T1 PixepolisDocument15 pagesNaturales T1 PixepolisTrizPas encore d'évaluation
- examenCIENCIASNATURALEScuartoBERTHA PDFDocument2 pagesexamenCIENCIASNATURALEScuartoBERTHA PDFSandy GalePas encore d'évaluation
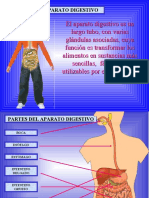
- Aparato Digestivo 40789 17450Document19 pagesAparato Digestivo 40789 17450Dany Villarroel MuñozPas encore d'évaluation
- MiologiaDocument4 pagesMiologiaalejandro terrazasPas encore d'évaluation
- Prefijos y Sufijos Por Aparatos y SistemasDocument26 pagesPrefijos y Sufijos Por Aparatos y Sistemasgelia100% (2)
- Guia Adhesion Celular - 4toELECTIVODocument4 pagesGuia Adhesion Celular - 4toELECTIVOMaria Alicia Bravo ArriagadaPas encore d'évaluation
- U1.LA Organización Cuerpo Humano1Document4 pagesU1.LA Organización Cuerpo Humano1GemmamocerPas encore d'évaluation
- El Sistema Digestivo para Niños para Primer Grado de PrimariaDocument5 pagesEl Sistema Digestivo para Niños para Primer Grado de PrimariaMau Castro80% (5)
- Cetaceos Sirenios Tortugas Guia Identificacion Caribe Pacifico ColombianoDocument126 pagesCetaceos Sirenios Tortugas Guia Identificacion Caribe Pacifico ColombianoGILMAR NIETOPas encore d'évaluation
- Tejido Animal - Biologia General - EP de Enfermeria 2020Document22 pagesTejido Animal - Biologia General - EP de Enfermeria 2020estehefaniPas encore d'évaluation
- La Digestion 5ºDocument4 pagesLa Digestion 5ºDARIO ZAMORAPas encore d'évaluation
- 4to Foro de Coccidia AVECAGDocument73 pages4to Foro de Coccidia AVECAGJesus Santos OrtizPas encore d'évaluation
- Aparato CirculatorioDocument7 pagesAparato CirculatorioNatita Garrido100% (1)
- Los Órganos ReproductoresDocument2 pagesLos Órganos Reproductoreslinder TaumaPas encore d'évaluation
- Guia 6 Ciencias Naturales Ciclo Tres PDFDocument4 pagesGuia 6 Ciencias Naturales Ciclo Tres PDFLuisa RuizPas encore d'évaluation
- CAPÍTULO 15 - Sistema CardiovascularDocument13 pagesCAPÍTULO 15 - Sistema CardiovascularVirginia Isabel Champney DubuissonPas encore d'évaluation
- Yupaykuna Los Numeros 8vo y 9noDocument5 pagesYupaykuna Los Numeros 8vo y 9noWILMER HUATATOCAPas encore d'évaluation
- Órganos Del Sistema LinfáticoDocument44 pagesÓrganos Del Sistema LinfáticoNathalySinchiPas encore d'évaluation
- Cuadro de Funciones de Los Seres VivosDocument9 pagesCuadro de Funciones de Los Seres VivosMARLON ALBERTO CASCO RAMIREZPas encore d'évaluation
- Deefensa AntidepredatoriaDocument22 pagesDeefensa AntidepredatoriaIndiana Valdez OvallePas encore d'évaluation
- Triptico Sis DigestivoDocument2 pagesTriptico Sis DigestivoBeatriz HinojosaPas encore d'évaluation
- La CelulaDocument3 pagesLa CelulaCristian LlanosPas encore d'évaluation
- Pene WikiDocument3 pagesPene WikiDragoniel PozoPas encore d'évaluation
- Datos Anatomicos Cuerpo HumanoDocument1 pageDatos Anatomicos Cuerpo Humanojose alfredo salazar huertaPas encore d'évaluation
- Actividad 1 Educacion Fisica (Santiago Guaqueta Rojas) 11-04Document6 pagesActividad 1 Educacion Fisica (Santiago Guaqueta Rojas) 11-04Santiago GuaquetaPas encore d'évaluation
- Actividades Unidad 5Document2 pagesActividades Unidad 5javier0% (2)
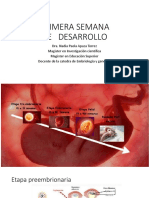
- Primera Semana de DesarrolloDocument221 pagesPrimera Semana de DesarrolloDaniela MujicaPas encore d'évaluation
- Guia Ciencias NaturalesDocument2 pagesGuia Ciencias Naturalesjessicapavez30Pas encore d'évaluation
- REPTILESDocument4 pagesREPTILESHumberto ReyesPas encore d'évaluation