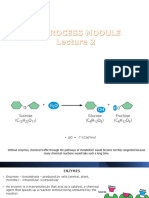
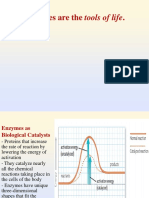
Vous aimerez peut-être aussi
- Production of NuggetDocument10 pagesProduction of NuggetNur AishaPas encore d'évaluation
- CrystallizationDocument18 pagesCrystallizationNur AishaPas encore d'évaluation
- New Approach To Heap Leach Modeling and Scale UpDocument18 pagesNew Approach To Heap Leach Modeling and Scale UpGeorgi SavovPas encore d'évaluation
- Primary Reformer TubesDocument10 pagesPrimary Reformer TubesAhmed ELmlahyPas encore d'évaluation
- Lecture 1 - The Kinetics of Enzyme Catalyzed ReactionsDocument18 pagesLecture 1 - The Kinetics of Enzyme Catalyzed ReactionsGenevive S. de VeraPas encore d'évaluation
- Enzyme Kinetics and Applications (Part 1a: Kinetics of Enzyme Catalyzed Reactions)Document26 pagesEnzyme Kinetics and Applications (Part 1a: Kinetics of Enzyme Catalyzed Reactions)Nur AishaPas encore d'évaluation
- Biochemistry 1: - Biochemistry of Amino Acids - Biochemistry of Proteins - Portrait of Allosteric ProteinDocument55 pagesBiochemistry 1: - Biochemistry of Amino Acids - Biochemistry of Proteins - Portrait of Allosteric ProteinHiba N IkhmyesPas encore d'évaluation
- Enzyme and Enzyme KineticsDocument61 pagesEnzyme and Enzyme KineticsNaiomiPas encore d'évaluation
- ENZYMESDocument31 pagesENZYMESjuunisai6Pas encore d'évaluation
- BIOCHEMISTRY LECTURE NotesDocument31 pagesBIOCHEMISTRY LECTURE NotesFREDRICK OUNDOPas encore d'évaluation
- Materi 2Document34 pagesMateri 2siti purnamaPas encore d'évaluation
- SCE3204 Lecture 2Document31 pagesSCE3204 Lecture 2ainomugisha arnoldPas encore d'évaluation
- Biochemistry - Enzyme-Protein Function and Metabolism 20180202Document78 pagesBiochemistry - Enzyme-Protein Function and Metabolism 20180202Artika Muliany TindaonPas encore d'évaluation
- Enzymes and Their RegulationDocument48 pagesEnzymes and Their RegulationChathur Chandrasoorya100% (1)
- Medical Biochemistry: Enzymes: Mr. Genaro F. Alderite JR, MsermDocument58 pagesMedical Biochemistry: Enzymes: Mr. Genaro F. Alderite JR, MsermNash DeniegaPas encore d'évaluation
- 4 EnzymesDocument33 pages4 EnzymesmjmonfortePas encore d'évaluation
- 12enzymes NewDocument39 pages12enzymes NewEman QawasmiPas encore d'évaluation
- Enzymes PresDocument34 pagesEnzymes PresRasmia SalamPas encore d'évaluation
- Lecture 2Document18 pagesLecture 2warden tambiPas encore d'évaluation
- Enzymes:: The Nature's CatalystsDocument51 pagesEnzymes:: The Nature's Catalystskrk100% (1)
- Enzyme: Ns. Shila Wisnasari, S.Kep., M.BiomedDocument55 pagesEnzyme: Ns. Shila Wisnasari, S.Kep., M.BiomedSyahdaJuvenilProfitamelaPas encore d'évaluation
- ERT211 CHP 1-2 PDFDocument27 pagesERT211 CHP 1-2 PDFsobiyamaragathavelPas encore d'évaluation
- 1 Enzyme Regulation in Biochemical PathwaysDocument29 pages1 Enzyme Regulation in Biochemical PathwayshadeelPas encore d'évaluation
- Enzymes & Energetics - CTPWDocument42 pagesEnzymes & Energetics - CTPWjessicatieuuPas encore d'évaluation
- Enzymes PresDocument57 pagesEnzymes Pres4tqpxmts7zPas encore d'évaluation
- Chapter 2: Diagnostic Enzymology: E+S Es E+PDocument67 pagesChapter 2: Diagnostic Enzymology: E+S Es E+PadamPas encore d'évaluation
- SEMESTER AWAL 2017/2018: Dasar-Dasar Biokimia EnzimDocument38 pagesSEMESTER AWAL 2017/2018: Dasar-Dasar Biokimia EnzimFachril ismailPas encore d'évaluation
- Pre-Lab 3: 1. What Is A Catalyst and A Catalysis Reaction?Document3 pagesPre-Lab 3: 1. What Is A Catalyst and A Catalysis Reaction?Trần NguyênPas encore d'évaluation
- Biochemistry: By: Angela Marie Ferrer BSN 2BDocument13 pagesBiochemistry: By: Angela Marie Ferrer BSN 2BNoemi Martinez FerrerPas encore d'évaluation
- EnzymesDocument3 pagesEnzymesArabela SimanganPas encore d'évaluation
- Biomolecules and Cells:: Mr. Derrick Banda MSC, BSCDocument69 pagesBiomolecules and Cells:: Mr. Derrick Banda MSC, BSCAmon Sangulube100% (1)
- EnzymesDocument41 pagesEnzymesaa9140697874Pas encore d'évaluation
- Enzymes PDFDocument36 pagesEnzymes PDFAltamashPas encore d'évaluation
- Enzymes: by Nura Malahayati THP-FP UnsriDocument71 pagesEnzymes: by Nura Malahayati THP-FP UnsriIkhsan muttaqinPas encore d'évaluation
- Enzymes: Anushree ChaturvediDocument39 pagesEnzymes: Anushree ChaturvediDeepal ShahPas encore d'évaluation
- Enzymes. RagaviDocument63 pagesEnzymes. Ragavikavi bharathiPas encore d'évaluation
- Title: Lecture (9) Enzymes Present By: Associate Prof - Ingy BadawyDocument58 pagesTitle: Lecture (9) Enzymes Present By: Associate Prof - Ingy BadawyAbdo ElsaidPas encore d'évaluation
- EnzymologyDocument15 pagesEnzymologyhkrybmzxfbxbwnpfhnPas encore d'évaluation
- Enzymes Lecture 1 2015animationDocument80 pagesEnzymes Lecture 1 2015animationSarfaras KKPas encore d'évaluation
- 4.0 CHPT 4 - Enzymes and The Regulation of Biochemical PathwaysDocument41 pages4.0 CHPT 4 - Enzymes and The Regulation of Biochemical PathwaysshinymaterialsPas encore d'évaluation
- Enzymes - Part IIDocument71 pagesEnzymes - Part IIBarış KaplanPas encore d'évaluation
- Nzymes: By: Mrs. Kalaivani Sathish. M. Pharm, Assistant Professor, Pims - PanipatDocument63 pagesNzymes: By: Mrs. Kalaivani Sathish. M. Pharm, Assistant Professor, Pims - Panipaturmila pandeyPas encore d'évaluation
- 02 EnzymesDocument63 pages02 EnzymesFatish BanguraPas encore d'évaluation
- 04 EnzymologyDocument39 pages04 EnzymologyFrances FranciscoPas encore d'évaluation
- 1380960835.1698lecture 4 & 5. EnzymesDocument30 pages1380960835.1698lecture 4 & 5. EnzymesKedir MohammedPas encore d'évaluation
- Module 2 Enzyme TransDocument11 pagesModule 2 Enzyme TransKathleen CedilloPas encore d'évaluation
- "Helper" Protein Molecules: EnzymesDocument7 pages"Helper" Protein Molecules: EnzymesMuhammad Junaid IqbalPas encore d'évaluation
- Classification of Enzymes: Sadia SayedDocument25 pagesClassification of Enzymes: Sadia SayedmiadelfiorPas encore d'évaluation
- Enzyme ChemistryDocument40 pagesEnzyme Chemistryakshif KhanPas encore d'évaluation
- Biological Molecules: Prepared By: Mrs. Eden C. SanchezDocument50 pagesBiological Molecules: Prepared By: Mrs. Eden C. SanchezBernard D. Fajardo Jr.100% (1)
- Enzyme UpdatedDocument42 pagesEnzyme UpdatedBukhari DiangkaPas encore d'évaluation
- Enzymes Dr. Madhur GuptaDocument89 pagesEnzymes Dr. Madhur GuptaAarohiPas encore d'évaluation
- EnzymesDocument5 pagesEnzymeseyosiyas tekleweyinPas encore d'évaluation
- EnzymesDocument46 pagesEnzymesHighlifePas encore d'évaluation
- LEC 06 - Enzyme MechanismsDocument70 pagesLEC 06 - Enzyme Mechanismseliza makPas encore d'évaluation
- CHP 4 BiocatalysisDocument47 pagesCHP 4 BiocatalysisSIMBA The Lion KingPas encore d'évaluation
- Factors Ppt5Document26 pagesFactors Ppt5DanishPas encore d'évaluation
- Enzymes PDFDocument50 pagesEnzymes PDFMuhammad Luqman NisaPas encore d'évaluation
- EnzymesDocument205 pagesEnzymes464464naifPas encore d'évaluation
- Enzyme Structure, Classification and Mechanism of ActionDocument35 pagesEnzyme Structure, Classification and Mechanism of ActionAnonymous cgKtuWzPas encore d'évaluation
- Introduction To Enzymology: - Functional Properties - Nomenclature - Enzyme Specificity - Enzyme RegulationDocument16 pagesIntroduction To Enzymology: - Functional Properties - Nomenclature - Enzyme Specificity - Enzyme RegulationYohandry Cruz AvilaPas encore d'évaluation
- Enzymes Are The Tools of LifeDocument31 pagesEnzymes Are The Tools of LifedrhydrogenPas encore d'évaluation
- Enzymes - BiochemistryDocument40 pagesEnzymes - Biochemistrysunil patelPas encore d'évaluation
- A-level Biology Revision: Cheeky Revision ShortcutsD'EverandA-level Biology Revision: Cheeky Revision ShortcutsÉvaluation : 5 sur 5 étoiles5/5 (5)
- Nguyen PDFDocument245 pagesNguyen PDFNur AishaPas encore d'évaluation
- Halal Expertise AsblDocument2 pagesHalal Expertise AsblNur AishaPas encore d'évaluation
- Quality Improvement of Tilapia Fish Nuggets by Addition of Legume Flour As ExtendersDocument13 pagesQuality Improvement of Tilapia Fish Nuggets by Addition of Legume Flour As ExtendersNur AishaPas encore d'évaluation
- ASSIGNMENT 1 HalalDocument6 pagesASSIGNMENT 1 HalalNur AishaPas encore d'évaluation
- COA ProceduresDocument20 pagesCOA ProceduresNur AishaPas encore d'évaluation
- Ethanol Fermentation in An Immobilized Cell Reactor Using Saccharomyces CerevisiaeDocument10 pagesEthanol Fermentation in An Immobilized Cell Reactor Using Saccharomyces CerevisiaeNur AishaPas encore d'évaluation
- Hazop Study.xDocument17 pagesHazop Study.xNur AishaPas encore d'évaluation
- Preparation of Media: Microbiology BIOL 275Document9 pagesPreparation of Media: Microbiology BIOL 275Nur AishaPas encore d'évaluation
- FermentationDocument8 pagesFermentationNur AishaPas encore d'évaluation
- Production of Bioenergy and Biochemicals From Industrial and Agricultural WastewaterDocument9 pagesProduction of Bioenergy and Biochemicals From Industrial and Agricultural WastewaterNur AishaPas encore d'évaluation
- Study On Production, Extraction and Analysis of Polyhydroxyalkanoate (PHA) From Bacterial IsolatesDocument8 pagesStudy On Production, Extraction and Analysis of Polyhydroxyalkanoate (PHA) From Bacterial IsolatesNur AishaPas encore d'évaluation
- ERT 319 Industrial Waste Treatment: Semester 1 2012/2013Document18 pagesERT 319 Industrial Waste Treatment: Semester 1 2012/2013Nur AishaPas encore d'évaluation
- Waste Minimization and ManagementDocument32 pagesWaste Minimization and ManagementNur AishaPas encore d'évaluation
- 5 Yield PDFDocument2 pages5 Yield PDFNur AishaPas encore d'évaluation
- Antroquinonol - Synthesis Process 29 April 2022Document10 pagesAntroquinonol - Synthesis Process 29 April 2022Prashant JadhavPas encore d'évaluation
- Metabolismo MicrobianoDocument165 pagesMetabolismo MicrobianoIsabella SanzPas encore d'évaluation
- Chapter 3: Oxidation and Reduction Form 5 Chemistry Title: Rusting As A Redox ReactionDocument3 pagesChapter 3: Oxidation and Reduction Form 5 Chemistry Title: Rusting As A Redox Reactionryder1man6433Pas encore d'évaluation
- Chemistry Answer Key Preboard 1Document15 pagesChemistry Answer Key Preboard 1Vibhas ReddyPas encore d'évaluation
- Catalogo SurvavierDocument24 pagesCatalogo SurvavierPhanor ReveloPas encore d'évaluation
- 3 Methyl 1 ButanolDocument2 pages3 Methyl 1 ButanolHamada HamadaPas encore d'évaluation
- CH 06Document34 pagesCH 06Firas Abu talebPas encore d'évaluation
- Preparation of SaltsDocument8 pagesPreparation of SaltsRose MusariraPas encore d'évaluation
- A Level Chemistry Core Practical 12 - TM ComplexDocument4 pagesA Level Chemistry Core Practical 12 - TM ComplexNamira ShameemPas encore d'évaluation
- Demo Table of Prediction Methods For Feed Oil and Biodiesel PropertiesDocument9 pagesDemo Table of Prediction Methods For Feed Oil and Biodiesel PropertiesJessicalba LouPas encore d'évaluation
- Pickering Emulsion Polymerization Kinetics of StyreneDocument44 pagesPickering Emulsion Polymerization Kinetics of StyreneAnonymous 9bc25fDPas encore d'évaluation
- Bellwork-Ionic & Metallic Venn DiagramDocument32 pagesBellwork-Ionic & Metallic Venn Diagramarissa noorPas encore d'évaluation
- Coating Galvanized: SteelDocument33 pagesCoating Galvanized: SteelJulian SihombingPas encore d'évaluation
- Bio3110 - Biochemistry Ii-Intermediary Metabolism Enzyme and Enzyme KineticsDocument11 pagesBio3110 - Biochemistry Ii-Intermediary Metabolism Enzyme and Enzyme KineticsNaiomiPas encore d'évaluation
- CaseinDocument11 pagesCaseinsudhuPas encore d'évaluation
- Electrons in Oxygen - and Sulphur-Containing Ligands PDFDocument132 pagesElectrons in Oxygen - and Sulphur-Containing Ligands PDFJose PerezPas encore d'évaluation
- CP-29 Carbopol Aqua SF-1 Product SummaryDocument2 pagesCP-29 Carbopol Aqua SF-1 Product SummaryadrianaPas encore d'évaluation
- LSST-2021-14974 Proof HiDocument21 pagesLSST-2021-14974 Proof HiDharmPalPas encore d'évaluation
- Equivalencia AceroDocument30 pagesEquivalencia AceroJose VelosaPas encore d'évaluation
- L1 Measurement-StudentDocument66 pagesL1 Measurement-StudentNN JKPas encore d'évaluation
- 9.1.1 Introduction To Oxidation and ReductionDocument24 pages9.1.1 Introduction To Oxidation and ReductionPatrick AbidraPas encore d'évaluation
- Effect of Boiled Cassava Leaves (Manihot Esculenta Crantz) On Total Phenolic, Flavonoid and Its Antioxidant ActivityDocument12 pagesEffect of Boiled Cassava Leaves (Manihot Esculenta Crantz) On Total Phenolic, Flavonoid and Its Antioxidant ActivityOxide HelixPas encore d'évaluation
- 11 Chemistry Eng SM 2024Document296 pages11 Chemistry Eng SM 2024Sumit YadavPas encore d'évaluation
- Ceb1013 - Lecture 8Document70 pagesCeb1013 - Lecture 8api-533764142Pas encore d'évaluation
- For Teachers Only P .S.-CH: Physical Setting/ChemistryDocument10 pagesFor Teachers Only P .S.-CH: Physical Setting/ChemistryAAVANIPas encore d'évaluation
- Rate of Flow of Iv FluidsDocument29 pagesRate of Flow of Iv Fluidsamethyst grande100% (1)
- 2021 Master CatalogDocument330 pages2021 Master Cataloggeetha raniPas encore d'évaluation
- GasTec PSA N2 Gen Systems PresentationDocument12 pagesGasTec PSA N2 Gen Systems Presentationdj22500Pas encore d'évaluation