Vous aimerez peut-être aussi
- Clase 01 - A Sem. Cardiovascular 2020-2 Historia Clinica. Sintomas Principales 1° ParteDocument50 pagesClase 01 - A Sem. Cardiovascular 2020-2 Historia Clinica. Sintomas Principales 1° ParteDimitry Antonov Tartarov PupanovPas encore d'évaluation
- Sistema Endocrino Historia Clinica 2019Document49 pagesSistema Endocrino Historia Clinica 2019Dimitry Antonov Tartarov PupanovPas encore d'évaluation
- Clase03 Auscultacion Cardiaca I-Ii PDFDocument25 pagesClase03 Auscultacion Cardiaca I-Ii PDFDimitry Antonov Tartarov PupanovPas encore d'évaluation
- Clase02 Semiologia CardioDocument97 pagesClase02 Semiologia CardioGoku Naruto VeguetaPas encore d'évaluation
- Diabetes DR LopezDocument64 pagesDiabetes DR LopezDimitry Antonov Tartarov PupanovPas encore d'évaluation
- Practica Semana 4 - Casos Clínicos PDFDocument27 pagesPractica Semana 4 - Casos Clínicos PDFDimitry Antonov Tartarov PupanovPas encore d'évaluation
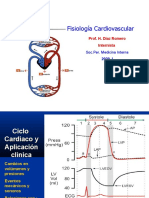
- Clase 2 Ciclo CardiacoDocument14 pagesClase 2 Ciclo CardiacoDimitry Antonov Tartarov PupanovPas encore d'évaluation
- Clase01-B Semiolocia CV 2020-2 Examen de Los Pulsos Art.Document25 pagesClase01-B Semiolocia CV 2020-2 Examen de Los Pulsos Art.Dimitry Antonov Tartarov PupanovPas encore d'évaluation
- Manual de ProblemasDocument160 pagesManual de ProblemasRakel Dreaming Inblue0% (1)
- Practica Semana 4 - Casos Clínicos PDFDocument27 pagesPractica Semana 4 - Casos Clínicos PDFDimitry Antonov Tartarov PupanovPas encore d'évaluation
- Arco Reflejo Placa MioneuralDocument26 pagesArco Reflejo Placa MioneuralDimitry Antonov Tartarov PupanovPas encore d'évaluation
- Diabetes TelesupDocument20 pagesDiabetes TelesupDimitry Antonov Tartarov PupanovPas encore d'évaluation
- Clase 3 HEMODINAMIA OkDocument28 pagesClase 3 HEMODINAMIA OkDimitry Antonov Tartarov PupanovPas encore d'évaluation
- Práctica N°9-Dr. Raúl Diaz - Matriz de Monitoreo, Supervisión y EvaluaciónDocument9 pagesPráctica N°9-Dr. Raúl Diaz - Matriz de Monitoreo, Supervisión y EvaluaciónDimitry Antonov Tartarov Pupanov100% (2)
- ESQUEMASDocument23 pagesESQUEMASDimitry Antonov Tartarov PupanovPas encore d'évaluation
- ESQUEMASDocument23 pagesESQUEMASDimitry Antonov Tartarov PupanovPas encore d'évaluation
- Supra R RenalDocument116 pagesSupra R RenalDimitry Antonov Tartarov PupanovPas encore d'évaluation
- Villapepas Ed. 2019 - Folleto 1Document4 pagesVillapepas Ed. 2019 - Folleto 1Dimitry Antonov Tartarov PupanovPas encore d'évaluation
- Anatomia y Patologia SuprarrenalDocument27 pagesAnatomia y Patologia SuprarrenalDimitry Antonov Tartarov PupanovPas encore d'évaluation
- Anatomia y Patologia de TiroidesDocument22 pagesAnatomia y Patologia de TiroidesDimitry Antonov Tartarov PupanovPas encore d'évaluation
- 6 TuberculosisDocument2 pages6 TuberculosisZoe Camila Alegre CastilloPas encore d'évaluation
- Radiologia SNCDocument52 pagesRadiologia SNCDimitry Antonov Tartarov PupanovPas encore d'évaluation
- Sindrome de Hipo Hiper Funcion Hipofisiaria 2019Document31 pagesSindrome de Hipo Hiper Funcion Hipofisiaria 2019Dimitry Antonov Tartarov PupanovPas encore d'évaluation
- Anatomia y Patologia de HipofisisDocument19 pagesAnatomia y Patologia de HipofisisDimitry Antonov Tartarov PupanovPas encore d'évaluation
- Cardio 2 - TelesupDocument73 pagesCardio 2 - TelesupDimitry Antonov Tartarov PupanovPas encore d'évaluation
- HC 34 Soma, CabezaDocument2 pagesHC 34 Soma, CabezaDimitry Antonov Tartarov PupanovPas encore d'évaluation
- HC 16 - Julián ArceDocument13 pagesHC 16 - Julián ArceDimitry Antonov Tartarov PupanovPas encore d'évaluation
- Análisis del líquido pleural: estudio citológico, bioquímico y microbiológicoDocument31 pagesAnálisis del líquido pleural: estudio citológico, bioquímico y microbiológicoDimitry Antonov Tartarov PupanovPas encore d'évaluation
- Temperatura Corporal Fiebre UrpDocument39 pagesTemperatura Corporal Fiebre UrpDimitry Antonov Tartarov PupanovPas encore d'évaluation
- Marcadores TumoralesDocument30 pagesMarcadores TumoralesDimitry Antonov Tartarov PupanovPas encore d'évaluation
- DocumentoDocument2 pagesDocumentopedorPas encore d'évaluation
- Lab 1 Medir Campo ElectricoDocument2 pagesLab 1 Medir Campo ElectricoSebastian MartinezPas encore d'évaluation
- Rúbrica para Evaluar Sistema Locomotor 2Document1 pageRúbrica para Evaluar Sistema Locomotor 2Muriel SotoPas encore d'évaluation
- Reporte Cedula Referencia Salud alumno-SEC216Document1 pageReporte Cedula Referencia Salud alumno-SEC216Julio OsornioPas encore d'évaluation
- Evidencia Semana3Document4 pagesEvidencia Semana3Yesid VasquezPas encore d'évaluation
- Instrucciones para El CALCULO DE METASDocument3 pagesInstrucciones para El CALCULO DE METASMarilse Sanchez Perez0% (1)
- BQ 96Document87 pagesBQ 96Meliza Chuquiyauri IrcañaupaPas encore d'évaluation
- Liberación de Hormigón.Document2 pagesLiberación de Hormigón.Franklin GarcíaPas encore d'évaluation
- PRQ 3219 Tema 8 PDFDocument9 pagesPRQ 3219 Tema 8 PDFAle SuaznabarPas encore d'évaluation
- Pequeña Planta Tratamiento Agua Potable TacnaDocument4 pagesPequeña Planta Tratamiento Agua Potable TacnaAlejandro Medina MendiaPas encore d'évaluation
- A MINUTADocument2 pagesA MINUTARomina Nuñez MoyaPas encore d'évaluation
- Informe Del SeminarioDocument10 pagesInforme Del SeminarioJeyson Dávila GuerreroPas encore d'évaluation
- 50.1 Componentes Del Costo de Los Inventarios (NIC 2) DUO PRACTICUMDocument88 pages50.1 Componentes Del Costo de Los Inventarios (NIC 2) DUO PRACTICUMEymiCristinaPas encore d'évaluation
- Juan M. Gonzalez - Concepto Ingeniería EconómicaDocument2 pagesJuan M. Gonzalez - Concepto Ingeniería Económicacpyin88Pas encore d'évaluation
- Mineria Artezanal Convencional e IlegalDocument16 pagesMineria Artezanal Convencional e IlegalAlexander Teves ZapanaPas encore d'évaluation
- ACTIVIDAD #3 - ABC - ComunicaciónDocument2 pagesACTIVIDAD #3 - ABC - ComunicaciónCasandra Joaquin VelasquezPas encore d'évaluation
- Velasco El Habitát de La Nobleza Siglo XviiiDocument42 pagesVelasco El Habitát de La Nobleza Siglo XviiiBetnaly González YáñezPas encore d'évaluation
- Estudio Ambiental - CondebambaDocument66 pagesEstudio Ambiental - CondebambaJorge Chuzon JimenezPas encore d'évaluation
- 2000 CastillaLaMancha ProblemaQuímicaDocument2 pages2000 CastillaLaMancha ProblemaQuímicaMaria LunesPas encore d'évaluation
- Tercer Parcial Con ClaveDocument4 pagesTercer Parcial Con ClaveJulissa Giselle Campos LópezPas encore d'évaluation
- BARRÍAvolcan Osorno 2014Document3 pagesBARRÍAvolcan Osorno 2014Andrea CastroPas encore d'évaluation
- CAPITULO 34 Antiinflamatorios, Antipiréticos y AnalgésicosDocument42 pagesCAPITULO 34 Antiinflamatorios, Antipiréticos y AnalgésicoswendypardoramirezPas encore d'évaluation
- Las 10 Ideas Principales Cap 18Document3 pagesLas 10 Ideas Principales Cap 18Lynda Greisy Luque CoaritePas encore d'évaluation
- Catalogo StarkerDocument24 pagesCatalogo Starkerdavekoz33% (3)
- Costos RelevantesDocument18 pagesCostos RelevantesYesenia Hallasi Ticona100% (1)
- Instalar Microsoft Dynamics CRM Server 2016Document11 pagesInstalar Microsoft Dynamics CRM Server 2016Jose A Aparicio FernandezPas encore d'évaluation
- 1 PruebaDocument20 pages1 PruebaRicPas encore d'évaluation
- Clase 4 TEORÍADocument4 pagesClase 4 TEORÍARos BarbozaPas encore d'évaluation
- Trabajo Escrito Sobre Motores - 4MM5Document9 pagesTrabajo Escrito Sobre Motores - 4MM5Jahzeel Alexis Arias CarreraPas encore d'évaluation
- Control Del Lectura 1 Idel Alberto Montero Ba-19-10522Document6 pagesControl Del Lectura 1 Idel Alberto Montero Ba-19-10522MUSICA URBANA, LATINA LETRASPas encore d'évaluation