Vous aimerez peut-être aussi
- 2 Taller Infecciones IntrahospitalariasDocument3 pages2 Taller Infecciones IntrahospitalariasCamiPas encore d'évaluation
- DigoxinaDocument2 pagesDigoxinaCami100% (1)
- Casos Clinicos Inmunosupresores y Anticuerpos MonoclonalesDocument3 pagesCasos Clinicos Inmunosupresores y Anticuerpos MonoclonalesCamiPas encore d'évaluation
- 11 Helmintos P1Document44 pages11 Helmintos P1CamiPas encore d'évaluation
- 10 Generalidades Parasitologia - Parásitos IntestinalesDocument43 pages10 Generalidades Parasitologia - Parásitos IntestinalesCamiPas encore d'évaluation
- 9 Generalidades VirusDocument55 pages9 Generalidades VirusCamiPas encore d'évaluation
- Nefropatia DiabeticaDocument10 pagesNefropatia DiabeticaCamiPas encore d'évaluation
- Desarrollo de La Placenta y Anexos EmbrionariosDocument93 pagesDesarrollo de La Placenta y Anexos EmbrionariosCami0% (1)
- Fisiología IntroduciónDocument70 pagesFisiología IntroduciónCamiPas encore d'évaluation
- Intoxicacion Por MercurioDocument8 pagesIntoxicacion Por MercurioCamiPas encore d'évaluation
- Wuolah-free-Tema 3. Tejido Conectivo - PDFDocument18 pagesWuolah-free-Tema 3. Tejido Conectivo - PDFCamiPas encore d'évaluation
- Tema 3. La Herencia CromosómicaDocument12 pagesTema 3. La Herencia Cromosómicamaripeco.2008Pas encore d'évaluation
- Ejercicio Señales 2 FisiologiaDocument3 pagesEjercicio Señales 2 FisiologiaYoali Asiain LeconaPas encore d'évaluation
- Estudio de Las Alteraciones en La Morfología EritrocitariaDocument65 pagesEstudio de Las Alteraciones en La Morfología Eritrocitariaquicksilver999Pas encore d'évaluation
- MFPH I Rep Mic Clave TeoricoDocument5 pagesMFPH I Rep Mic Clave Teoricob bnnmmmn bnnbmbm100% (2)
- Trabajo de BiologíaDocument4 pagesTrabajo de BiologíaCristofer GuamboPas encore d'évaluation
- Importancia de La Ecologia en La MedicinaDocument4 pagesImportancia de La Ecologia en La MedicinaRicardo SanchezPas encore d'évaluation
- Cuadro Sinoptico de Medio de CultivoDocument2 pagesCuadro Sinoptico de Medio de Cultivorommel alfonso bolainaPas encore d'évaluation
- Biodiversidad y Sociodiversida1Document12 pagesBiodiversidad y Sociodiversida1Alexandra Giraldo PerezPas encore d'évaluation
- Telomeros y Molecula de Agua - Buscar Con GoogleDocument2 pagesTelomeros y Molecula de Agua - Buscar Con GoogleYamil Ekel MilgronPas encore d'évaluation
- Introducción SISTEMA OSEODocument14 pagesIntroducción SISTEMA OSEOPaula JimenezPas encore d'évaluation
- Microcurriculo Del Curso Trabajo SocialDocument11 pagesMicrocurriculo Del Curso Trabajo SocialAngie SernaPas encore d'évaluation
- Cuestionario Célula Quinto GradoDocument1 pageCuestionario Célula Quinto GradoLADY CARVAJALINOPas encore d'évaluation
- Informe 2 Lab GeneticsDocument12 pagesInforme 2 Lab GeneticsNorma100% (2)
- Bases Neurobiologicas de La ConductaDocument23 pagesBases Neurobiologicas de La ConductaUSMP FN ARCHIVOS100% (1)
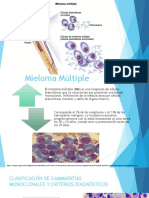
- Mieloma MúltipleDocument20 pagesMieloma MúltipleCarlos Benitez100% (1)
- Grupo 4 Proyecto Fin de CicloDocument6 pagesGrupo 4 Proyecto Fin de CicloBianca Yubraxis Saavedra GonzalezPas encore d'évaluation
- Vacuna Oral Rabia Dra. Manolo Fernandez FARVETDocument32 pagesVacuna Oral Rabia Dra. Manolo Fernandez FARVETstefhany valdeiglesiasPas encore d'évaluation
- Anticuerpos Monoclonales y Policlonales 2017Document21 pagesAnticuerpos Monoclonales y Policlonales 2017AdriPerezPas encore d'évaluation
- Unidad II Células, Organos y Tejidos Del Sistema InmuneDocument48 pagesUnidad II Células, Organos y Tejidos Del Sistema InmuneSam Son Soleil100% (3)
- Inmunología - Seminario - FinalDocument57 pagesInmunología - Seminario - FinalJEFFERSON ALBERTO RUIZ CHAUCAPas encore d'évaluation
- Etica Mapa MentalDocument1 pageEtica Mapa MentalNikoll Sofia Herrera Angarita0% (1)
- Etapa 2 - Taller Genetica Mendeliana GRUPO 30162A-954Document21 pagesEtapa 2 - Taller Genetica Mendeliana GRUPO 30162A-954Alexandra FlorezPas encore d'évaluation
- Breve Historia de La InmunologíaDocument2 pagesBreve Historia de La InmunologíaLina SuescunPas encore d'évaluation
- Anatomia Cpu Unprg Sistema Digestivo Cap-Ix PDFDocument10 pagesAnatomia Cpu Unprg Sistema Digestivo Cap-Ix PDFMiguel Ángel Zúñiga50% (2)
- Tema 11. La Célula ProcariotaDocument6 pagesTema 11. La Célula ProcariotaCristina Agudelo RamírezPas encore d'évaluation
- Informe 4 Bioquímica Seminario - Grupo 07Document12 pagesInforme 4 Bioquímica Seminario - Grupo 07Emery Sofía C. D. Bustamante0% (1)
- Comunicación Celular y Contracción MuscularDocument11 pagesComunicación Celular y Contracción MuscularCamila AndradePas encore d'évaluation
- Herencia MitocondrialDocument4 pagesHerencia MitocondrialLucas Christopher100% (1)
- Bioseguridad Expo I InyectoterapiaDocument16 pagesBioseguridad Expo I InyectoterapiaxxmalenaxxPas encore d'évaluation
- Bioquimica Prodeunp Sullana Unidad 1 BiotecnologiaDocument114 pagesBioquimica Prodeunp Sullana Unidad 1 BiotecnologiaRicardo Noé Ágreda PalominoPas encore d'évaluation