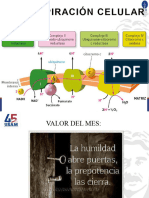
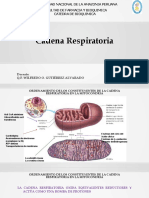
Vous aimerez peut-être aussi
- Cadena Respiratoria Fosforilacion OxidatDocument95 pagesCadena Respiratoria Fosforilacion OxidatMiguel Soni CasillasPas encore d'évaluation
- Cadena Respiratoria Ciclo 1-2020Document39 pagesCadena Respiratoria Ciclo 1-2020Grace AlvaradoPas encore d'évaluation
- Cadena Respiratoria y Fosforilación OxidativaDocument6 pagesCadena Respiratoria y Fosforilación OxidativaJosé GuamánPas encore d'évaluation
- Infografía de Guía de Cadena Estilo Minimalista en Colores SuavesDocument1 pageInfografía de Guía de Cadena Estilo Minimalista en Colores SuavesDiego Armando PerezPas encore d'évaluation
- Universidad Autónoma de Baja California Unidad Valle de Las Palmas Facultad de Ciencias de La Salud MedicinaDocument6 pagesUniversidad Autónoma de Baja California Unidad Valle de Las Palmas Facultad de Ciencias de La Salud MedicinaCesar Alexis Jiménez BasurtoPas encore d'évaluation
- Cadena RespiratoriaDocument26 pagesCadena Respiratoriales12michuPas encore d'évaluation
- Transporte Electrónico y Fosforilación OxidativaDocument33 pagesTransporte Electrónico y Fosforilación OxidativaJimmy ArroligaPas encore d'évaluation
- Tema 1 Introduccion Al Metabolismo y Señalización Celular 2018Document65 pagesTema 1 Introduccion Al Metabolismo y Señalización Celular 2018uniautonoma53Pas encore d'évaluation
- Cadena RespiratoriaDocument25 pagesCadena RespiratoriaKruzkaya MachicadoPas encore d'évaluation
- Transporte Electronico y Fosforilacion OxidativaDocument41 pagesTransporte Electronico y Fosforilacion OxidativanicolehurtadosPas encore d'évaluation
- Respiración CelularDocument46 pagesRespiración CelularRamses DeelPas encore d'évaluation
- Cadena Respiratoria y Fosforilación OxidativaDocument21 pagesCadena Respiratoria y Fosforilación OxidativaItalia BaezPas encore d'évaluation
- DG4 Presentacion Grupo3aDocument31 pagesDG4 Presentacion Grupo3aJorge SalomonPas encore d'évaluation
- Cadena RespiratoriaDocument1 pageCadena RespiratoriaJohann LópezPas encore d'évaluation
- Conferencia 17 La Fosforilación OxidativaDocument33 pagesConferencia 17 La Fosforilación OxidativaJoaquim JosephPas encore d'évaluation
- Fosforilación IDocument64 pagesFosforilación IYlaresPas encore d'évaluation
- MitocondriaDocument38 pagesMitocondriakremenchutzkyPas encore d'évaluation
- La Mitocondria Como Fuente de Energía eDocument23 pagesLa Mitocondria Como Fuente de Energía eAlejandra RojasPas encore d'évaluation
- Molécula de ATPDocument2 pagesMolécula de ATPmarianaioro13Pas encore d'évaluation
- Informe #3 - LaboratorioDocument12 pagesInforme #3 - LaboratorioJAVIER ANTHONY BUSTOS LOPEZPas encore d'évaluation
- Curvas de Concentraci N de Ox Geno y Concentraci N de Atp en Funci N Del Tiempo 1Document22 pagesCurvas de Concentraci N de Ox Geno y Concentraci N de Atp en Funci N Del Tiempo 1docsamu78Pas encore d'évaluation
- P7. Efecto de Los Inhibidores en La Cadena respiratoria-PROTOCOLODocument38 pagesP7. Efecto de Los Inhibidores en La Cadena respiratoria-PROTOCOLOJuan Rodríguez CarbonellPas encore d'évaluation
- Discu5 Fosforilacion OxidativaDocument9 pagesDiscu5 Fosforilacion OxidativaJosé LuisPas encore d'évaluation
- Unidad Ii Fosforilacion y Cadena RespiratoriaDocument65 pagesUnidad Ii Fosforilacion y Cadena Respiratoriaevelyna.gomezPas encore d'évaluation
- T 16 Cadena RespiratoriaDocument23 pagesT 16 Cadena RespiratoriaElias Del aguila PerezPas encore d'évaluation
- Dadores Aceptores y P/ODocument36 pagesDadores Aceptores y P/OFrancisco Abanto33% (3)
- Cadena RespiratoriaDocument27 pagesCadena RespiratoriaClaribel ValdezPas encore d'évaluation
- ACTIVIDAD PRACTICA N°6 - MITOCONDRIAS-PEROXISOMAS - Semana9Document5 pagesACTIVIDAD PRACTICA N°6 - MITOCONDRIAS-PEROXISOMAS - Semana9Laura MoyaPas encore d'évaluation
- Cadena RespiratoriaDocument4 pagesCadena RespiratoriaJoiss Chirinos100% (1)
- Fosforilación OxidativaDocument10 pagesFosforilación OxidativaPablo CaberoPas encore d'évaluation
- Tema 2 Conversión EnergéticaDocument48 pagesTema 2 Conversión Energéticamarcosh777Pas encore d'évaluation
- 3 Discusión BioquímicaDocument16 pages3 Discusión BioquímicaMilver EscalantePas encore d'évaluation
- Discusion 3Document17 pagesDiscusion 3Milver EscalantePas encore d'évaluation
- TEORICO Cadena Resp, Fosfor OxidativaDocument12 pagesTEORICO Cadena Resp, Fosfor OxidativaSofia CoriaPas encore d'évaluation
- Fosforilacion OxidativaDocument6 pagesFosforilacion Oxidativalore05Pas encore d'évaluation
- 3 Cadena Respiratoria y Fosforilacion OxidativaDocument12 pages3 Cadena Respiratoria y Fosforilacion OxidativaAngelina MerinoPas encore d'évaluation
- Organelos Productores de EnergíaDocument28 pagesOrganelos Productores de EnergíaKenia DayamPas encore d'évaluation
- Discusion 4 Bioca IDocument22 pagesDiscusion 4 Bioca Iestradaf6Pas encore d'évaluation
- Cadena y FosforilacionDocument35 pagesCadena y FosforilacionJulio SánchezPas encore d'évaluation
- T7. Cadena Respiratoria-CLAVDocument53 pagesT7. Cadena Respiratoria-CLAVPaolo Gianfranco Luna Victoria GutierrezPas encore d'évaluation
- CAPÍTULO 13. La Cadena Respiratoria y Fosforilación Oxidativa.Document11 pagesCAPÍTULO 13. La Cadena Respiratoria y Fosforilación Oxidativa.YunierValderramaPas encore d'évaluation
- Respiración CelularDocument23 pagesRespiración CelularAntonio Torres PeñaPas encore d'évaluation
- Clase ModeloDocument5 pagesClase ModeloFranklin Miñope MechánPas encore d'évaluation
- Teórico - Cadena de Transporte de Electrones. Fosforilación Oxidativa - Texto - 2023Document24 pagesTeórico - Cadena de Transporte de Electrones. Fosforilación Oxidativa - Texto - 2023Gustavo LopezPas encore d'évaluation
- Taller Metabolismo de CarbohidratosDocument3 pagesTaller Metabolismo de Carbohidratosbetincarlos3010100% (2)
- Energia Metabolica: Generación Y MantenimientoDocument23 pagesEnergia Metabolica: Generación Y MantenimientoSonia CostaPas encore d'évaluation
- 1 Transporte de Electrones LicenciaturaDocument32 pages1 Transporte de Electrones LicenciaturaGustavo LVPas encore d'évaluation
- Cadena Respiratoria y Fosforilación OxidativaDocument18 pagesCadena Respiratoria y Fosforilación OxidativaSantimioPas encore d'évaluation
- RESPIRACIÓNDocument53 pagesRESPIRACIÓNCarolina QuinteroPas encore d'évaluation
- Metabolismo IntermediarioDocument22 pagesMetabolismo Intermediariokarina Honorio vega100% (1)
- Preguntas RespiraciónDocument5 pagesPreguntas RespiraciónJuárez Chapoñan JeseniaPas encore d'évaluation
- Lourdes TripticoDocument2 pagesLourdes TripticoZul Ysminda Lorena Quispe50% (4)
- 3-2022 Cte y Fosforilacion OxidativaDocument56 pages3-2022 Cte y Fosforilacion OxidativaAbisai FuentesPas encore d'évaluation
- Cadena Respiratoria Exposicion FinalDocument28 pagesCadena Respiratoria Exposicion Finalesneider alexander perilla vergaraPas encore d'évaluation
- Resumen Unidad I y II - MetabólicaDocument7 pagesResumen Unidad I y II - MetabólicamarPas encore d'évaluation
- Tema 3. Ciclo Del Ácido CítricoDocument15 pagesTema 3. Ciclo Del Ácido CítricoPaola Ojeda FernándezPas encore d'évaluation
- Acceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012D'EverandAcceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012Pas encore d'évaluation
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017D'EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Pas encore d'évaluation
- El Estudio de Los Axiomas Matemáticos Es Propio de La FilosofíaDocument2 pagesEl Estudio de Los Axiomas Matemáticos Es Propio de La FilosofíaWalterSandovalPas encore d'évaluation
- Vasculogénesis y AngiogénesisDocument1 pageVasculogénesis y AngiogénesisWalterSandovalPas encore d'évaluation
- Anatomia Semana 14Document25 pagesAnatomia Semana 14WalterSandovalPas encore d'évaluation
- Study 1.1Document2 pagesStudy 1.1WalterSandovalPas encore d'évaluation
- Terapia GenicaDocument10 pagesTerapia GenicaWalterSandovalPas encore d'évaluation
- TerrorismoDocument8 pagesTerrorismoWalterSandovalPas encore d'évaluation
- TerrorismoDocument8 pagesTerrorismoWalterSandovalPas encore d'évaluation
- Ejercicios Quimica OrganicaDocument7 pagesEjercicios Quimica OrganicaCaro CruzPas encore d'évaluation
- Molienda y TamizadoDocument16 pagesMolienda y TamizadoVanessa CastroPas encore d'évaluation
- Preparación de Soluciones Estiven NewDocument5 pagesPreparación de Soluciones Estiven NewestivPas encore d'évaluation
- Apuntes UNIPRO 2022Document9 pagesApuntes UNIPRO 2022IMPRESION EZEIZAPas encore d'évaluation
- Anexo No. 2.2 Especificaciones Técnicas A.A Piso 4 Salitre.Document22 pagesAnexo No. 2.2 Especificaciones Técnicas A.A Piso 4 Salitre.NiltonGZPas encore d'évaluation
- Técnicas de Destilación y ExtracciónDocument6 pagesTécnicas de Destilación y ExtracciónJacko MistePas encore d'évaluation
- Investigacion Propiedades de Los MaterialesDocument17 pagesInvestigacion Propiedades de Los MaterialesJuanPas encore d'évaluation
- Reaccion de Los Hidrocarburos AromaticosDocument10 pagesReaccion de Los Hidrocarburos AromaticosJesuita mcPas encore d'évaluation
- HidrocarburosDocument8 pagesHidrocarburosCésar Vásquez DonetPas encore d'évaluation
- 07 Taller Reacciones Químicas y BalanceoDocument2 pages07 Taller Reacciones Químicas y BalanceoJan Carlos Digregory WolfPas encore d'évaluation
- Fosforitas TachiraDocument22 pagesFosforitas Tachiraelmoro2014Pas encore d'évaluation
- RE-10-LAB-018 QUIMICA I v7Document59 pagesRE-10-LAB-018 QUIMICA I v7renepiza95Pas encore d'évaluation
- Chocolate de MesaDocument5 pagesChocolate de MesaCesar Ivan Cañas SerratoPas encore d'évaluation
- Q1 Unidad5 Clase13 ReaccionesQuímicasDocument18 pagesQ1 Unidad5 Clase13 ReaccionesQuímicasbrayan david Lliuya SaldañaPas encore d'évaluation
- Comparación Entre AAS & ICP OESDocument8 pagesComparación Entre AAS & ICP OESporfiriosegundoPas encore d'évaluation
- Ejercicio 2 - SXDocument2 pagesEjercicio 2 - SXHayden IvanovicPas encore d'évaluation
- Preparacion de SuperficiesDocument13 pagesPreparacion de SuperficiesNelson Elias Ordenes CerdaPas encore d'évaluation
- Diagrama de Flujo FDF-1Document3 pagesDiagrama de Flujo FDF-1Mario David Moncayo OrtizPas encore d'évaluation
- Informe 4 ExtraccionDocument19 pagesInforme 4 ExtraccionMarielit Sharid Arredondo QuirozPas encore d'évaluation
- Yeso CartònDocument16 pagesYeso CartònMatias MontecinosPas encore d'évaluation
- 5° A Examen Periodo 3 - 2023 - RespuestasDocument9 pages5° A Examen Periodo 3 - 2023 - RespuestasIsai Pérez MartínezPas encore d'évaluation
- Cinetica Quimica TeoriaDocument143 pagesCinetica Quimica TeoriaAlexa Ceballos SalgadoPas encore d'évaluation
- Enlaces QuímicosDocument2 pagesEnlaces QuímicosRicardo Andrés Gómez AlbaPas encore d'évaluation
- Persulfato de Amonio-Hds2017Document10 pagesPersulfato de Amonio-Hds2017Exequiel Viera ZepedaPas encore d'évaluation
- Procedimiento para El ColorDocument16 pagesProcedimiento para El ColorDICKSON J JIMENEZ RUIZPas encore d'évaluation
- El NaSH en El Circuito de Flotacion Selectiva de MolibdenoDocument2 pagesEl NaSH en El Circuito de Flotacion Selectiva de MolibdenoSISQUIM - Luis MejíasPas encore d'évaluation
- Anfigenos Practica N°11Document19 pagesAnfigenos Practica N°11Jhois Calderon MontesPas encore d'évaluation
- 3.1. Concepto e Importancia de La Absorción.Document5 pages3.1. Concepto e Importancia de La Absorción.Guadalupe Campos100% (1)
- Resina Colada EGDocument1 pageResina Colada EGariztiaPas encore d'évaluation
- Clase 3 Menbrana Plasmatica y DifusionDocument29 pagesClase 3 Menbrana Plasmatica y DifusionNancy GarofaloPas encore d'évaluation