Vous aimerez peut-être aussi
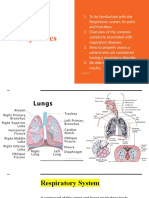
- Respiratory Physiology NoteDocument19 pagesRespiratory Physiology NotePeter Wong0% (1)
- Atelectasis, (Lung Collapse) A Simple Guide To The Condition, Diagnosis, Treatment And Related DiseasesD'EverandAtelectasis, (Lung Collapse) A Simple Guide To The Condition, Diagnosis, Treatment And Related DiseasesPas encore d'évaluation
- Cardio NoteDocument17 pagesCardio NoteMustak AhmedPas encore d'évaluation
- Respiratory PhysiologyDocument30 pagesRespiratory PhysiologyBarbara Sakura RiawanPas encore d'évaluation
- Lung Diseases, A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsD'EverandLung Diseases, A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsPas encore d'évaluation
- Respiratory Physiology Summary NotesDocument63 pagesRespiratory Physiology Summary NotesAlfredII100% (1)
- Gaseous ExchangeDocument37 pagesGaseous ExchangeSolángé MirembePas encore d'évaluation
- Problems With OxygenationDocument92 pagesProblems With OxygenationEbiPas encore d'évaluation
- Chapter 12Document28 pagesChapter 12براءة أحمد السلاماتPas encore d'évaluation
- M132 Respiratory 2022Document28 pagesM132 Respiratory 2022Bana ItanPas encore d'évaluation
- 12.respiratory SystemDocument52 pages12.respiratory SystemRulani PrincePas encore d'évaluation
- Respiratory SystemDocument32 pagesRespiratory SystemvhieeelgbmPas encore d'évaluation
- Respiration - VentilationDocument102 pagesRespiration - VentilationSodeinde SimeonPas encore d'évaluation
- Bio Notes (B3.1) Tuesday 30TH JanuaryDocument11 pagesBio Notes (B3.1) Tuesday 30TH JanuaryNeema Sarah MUTWIRIPas encore d'évaluation
- Compliance of LungsDocument37 pagesCompliance of LungsJasir KhanPas encore d'évaluation
- Ventilation and Respiratory VolumesDocument10 pagesVentilation and Respiratory Volumesalthea jade villadongaPas encore d'évaluation
- Diffusion and Human Respiratory SystemDocument9 pagesDiffusion and Human Respiratory SystemPriscilla AshwiniPas encore d'évaluation
- Biofluid Mechanics Chapter 3Document35 pagesBiofluid Mechanics Chapter 3AbcdPas encore d'évaluation
- Dr. Sunita Saxena - Breathing and Exchange of GassesDocument48 pagesDr. Sunita Saxena - Breathing and Exchange of GassesDivya AgarawalPas encore d'évaluation
- Fundamentalof Ventilation & PacemakerDocument22 pagesFundamentalof Ventilation & PacemakerLucila Lugo100% (1)
- Respiratory PhysiologyDocument98 pagesRespiratory PhysiologySurya SuryaPas encore d'évaluation
- RespirationDocument32 pagesRespirationAndersonPas encore d'évaluation
- Respiratory System: M.AzhariDocument142 pagesRespiratory System: M.AzhariLili ManaoPas encore d'évaluation
- Respiratory SystemDocument16 pagesRespiratory Systemapi-277775953Pas encore d'évaluation
- Basics of Respiratory Function: Mee Wah NG RSO Dubai 2002Document43 pagesBasics of Respiratory Function: Mee Wah NG RSO Dubai 2002chanlalPas encore d'évaluation
- Exchange of Gases Between The Atmosphere & The LungsDocument21 pagesExchange of Gases Between The Atmosphere & The LungsJulee Pearl GangosoPas encore d'évaluation
- Respiratory SystemDocument43 pagesRespiratory SystemAbuhuraira RafiqPas encore d'évaluation
- Chapter 11 - Respiratory SystemsDocument122 pagesChapter 11 - Respiratory Systemsnokate konkoorPas encore d'évaluation
- Respiration SystemDocument15 pagesRespiration Systemapi-283243804Pas encore d'évaluation
- Respiratory System-1Document84 pagesRespiratory System-1pranjal raiPas encore d'évaluation
- Chapter 22Document12 pagesChapter 22Jenn ParksPas encore d'évaluation
- Module 2Document40 pagesModule 2Sharanya BasakPas encore d'évaluation
- Breathing and Exchange of Gases-HumansDocument14 pagesBreathing and Exchange of Gases-Humansnihalbolla2023Pas encore d'évaluation
- BIO 2701 Lecture Notes Exchange With The EnvironmentDocument11 pagesBIO 2701 Lecture Notes Exchange With The EnvironmentDorothy MayakaPas encore d'évaluation
- L - 1 Physiology of Respiration IDocument25 pagesL - 1 Physiology of Respiration Ikaukab azimPas encore d'évaluation
- Respiratory Disorders Case 1 AsthmaDocument21 pagesRespiratory Disorders Case 1 AsthmajoatasouzaPas encore d'évaluation
- Nasal Cavity Nostrils Pharynx AIRDocument4 pagesNasal Cavity Nostrils Pharynx AIRDarwin CauilanPas encore d'évaluation
- Lecture Notes On Respiratory Physiology PDFDocument33 pagesLecture Notes On Respiratory Physiology PDFMiles HuiPas encore d'évaluation
- The Respiratory SystemDocument2 pagesThe Respiratory SystemconnievelardePas encore d'évaluation
- Respiratory System: Departement of Physiology Medical Faculty of Universitas Sumatera UtaraDocument142 pagesRespiratory System: Departement of Physiology Medical Faculty of Universitas Sumatera Utara292Vien HardiyantiPas encore d'évaluation
- 1.4 Lungs & DiseasesDocument10 pages1.4 Lungs & DiseasesRayonesh RayanaPas encore d'évaluation
- The Human Respiratory SystemDocument5 pagesThe Human Respiratory SystemnekromadavidPas encore d'évaluation
- Respiratory SystemDocument12 pagesRespiratory Systemab4943409Pas encore d'évaluation
- PhysioDocument72 pagesPhysioEINSTEIN2DPas encore d'évaluation
- The Mechanics of Breathing: Mashmum Shababa ShahidDocument31 pagesThe Mechanics of Breathing: Mashmum Shababa ShahidSHANJIDA ALI RIAPas encore d'évaluation
- Anatomy Review RespiratoryDocument8 pagesAnatomy Review RespiratoryHanami AsriPas encore d'évaluation
- L - 1 Physiology of Respiration IDocument28 pagesL - 1 Physiology of Respiration IshahabsworldPas encore d'évaluation
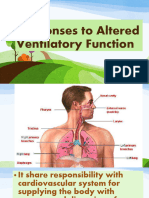
- NCM 118Responses-to-Altered-Ventilatory-FunctionDocument169 pagesNCM 118Responses-to-Altered-Ventilatory-FunctionAddah, Dhenaraiza H.Pas encore d'évaluation
- University of Guyana School of Medicine Med 1106 - Physiology I DR Kalima ThompsonDocument66 pagesUniversity of Guyana School of Medicine Med 1106 - Physiology I DR Kalima ThompsonKPas encore d'évaluation
- CP4. Respiratory SystemDocument28 pagesCP4. Respiratory Systemdafabc50Pas encore d'évaluation
- RESPIRATORY PhysiologyDocument101 pagesRESPIRATORY PhysiologyKhant Si ThuPas encore d'évaluation
- Respiratory Physiology ´ Respiratory Failure: Dr. Priyanka Gupta Dept of Anaesthesia AIIMS, RishikeshDocument100 pagesRespiratory Physiology ´ Respiratory Failure: Dr. Priyanka Gupta Dept of Anaesthesia AIIMS, RishikeshRinkiPas encore d'évaluation
- Anatomy & PhysiologyDocument42 pagesAnatomy & PhysiologyArah Lyn ApiagPas encore d'évaluation
- Dyamics of Respiration Composition VentilationDocument30 pagesDyamics of Respiration Composition VentilationUsama SajidPas encore d'évaluation
- Respiratory SystemDocument24 pagesRespiratory SystemArjunPas encore d'évaluation
- Hole'S Human Anatomy and Physiology: Powerpoint Lecture Outlines To AccompanyDocument52 pagesHole'S Human Anatomy and Physiology: Powerpoint Lecture Outlines To AccompanyEllen LiberacePas encore d'évaluation
- Breathing and Exchange of GasesDocument8 pagesBreathing and Exchange of GasesgoswamiuwuPas encore d'évaluation
- Gas Exchange in The LungsDocument30 pagesGas Exchange in The LungsYasasviPas encore d'évaluation
- How Do Humans Breathe? Science Book Age 8 | Children's Biology BooksD'EverandHow Do Humans Breathe? Science Book Age 8 | Children's Biology BooksPas encore d'évaluation
- Introduction - To - Physiology Slides of Doctor Mustafa ShehabatDocument30 pagesIntroduction - To - Physiology Slides of Doctor Mustafa ShehabatAmmar SmadiPas encore d'évaluation
- Homeostasis: Body Systems Maintain HomeostasisDocument46 pagesHomeostasis: Body Systems Maintain HomeostasisAmmar SmadiPas encore d'évaluation
- Dentistry Stydent (Physiology) Second Year - by Hashim Gazo - Slide # 1Document17 pagesDentistry Stydent (Physiology) Second Year - by Hashim Gazo - Slide # 1Hashim GhazoPas encore d'évaluation
- Chapter 12Document28 pagesChapter 12Ammar SmadiPas encore d'évaluation
- Principles of Neural and Hormonal CommunicationDocument25 pagesPrinciples of Neural and Hormonal CommunicationAmmar SmadiPas encore d'évaluation
- Body FluidsDocument59 pagesBody FluidsAmmar SmadiPas encore d'évaluation
- Homeostasis: Body Systems Maintain HomeostasisDocument46 pagesHomeostasis: Body Systems Maintain HomeostasisAmmar SmadiPas encore d'évaluation