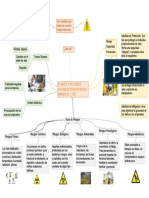
Vous aimerez peut-être aussi
- Cadena de Custodia de Muestras de Agua - TAR-01Document39 pagesCadena de Custodia de Muestras de Agua - TAR-01Gerardo Mendez Lopez100% (5)
- Seguridad e Higiene LaboralDocument5 pagesSeguridad e Higiene Laboralrosa luz zamoraPas encore d'évaluation
- Examen Parcial - Semana 4 - )Document13 pagesExamen Parcial - Semana 4 - )patoPas encore d'évaluation
- Expediente Técnico Anexo IDocument5 pagesExpediente Técnico Anexo IkarenPas encore d'évaluation
- INFORME #104 Certificacion Pptaria CMANDocument2 pagesINFORME #104 Certificacion Pptaria CMANVictor Reyes AntonioPas encore d'évaluation
- Accidentes Más Comunes en La Cocina ProfesionalDocument4 pagesAccidentes Más Comunes en La Cocina Profesionalroberto diazPas encore d'évaluation
- Arbol de ObjetivosDocument1 pageArbol de ObjetivosEduardoGuayguaPas encore d'évaluation
- Tesis FinalDocument37 pagesTesis FinalCarlos MeraPas encore d'évaluation
- Epidemiología General y Demografía SanitariaDocument13 pagesEpidemiología General y Demografía Sanitariaajss8838156Pas encore d'évaluation
- Espacio Retroperitoneal (Bouchet - 1978)Document10 pagesEspacio Retroperitoneal (Bouchet - 1978)Ingrid Alvarez Alvarado100% (1)
- Analisis de Las Obligaciones y Prohibiciones Del Trababador y PatronoDocument7 pagesAnalisis de Las Obligaciones y Prohibiciones Del Trababador y PatronoNazareth GarcíaPas encore d'évaluation
- Modelos de Evoluciones ClínicasDocument3 pagesModelos de Evoluciones ClínicasPaula QPas encore d'évaluation
- La Mujer Mariposa AmazonDocument456 pagesLa Mujer Mariposa AmazonManuel De La Torre Macias100% (1)
- Guia ADACAP v3 11 PDFDocument19 pagesGuia ADACAP v3 11 PDFAngel BautistaPas encore d'évaluation
- Enfermedades Óseas MetabólicasDocument23 pagesEnfermedades Óseas MetabólicasKevin Jose Huamani Torres100% (1)
- Cat Gear Oil 80W-90Document11 pagesCat Gear Oil 80W-90Carlos Manuel Basurto Hernandez100% (1)
- Cabrera Ramirez Ana Nelva Certificado Programa AuroraDocument1 pageCabrera Ramirez Ana Nelva Certificado Programa AuroraIvan CarreroPas encore d'évaluation
- Mapa Mental AmbienteDocument1 pageMapa Mental AmbienteDavidDanielRezaLopezPas encore d'évaluation
- Campamento Seguro 1Document8 pagesCampamento Seguro 1Valeria de Frias89% (9)
- EO-SST-PRO-00117-ANX A - Matriz IdentificacionADocument12 pagesEO-SST-PRO-00117-ANX A - Matriz IdentificacionAMobec Sa FetyPas encore d'évaluation
- Alimentos TransgenicosDocument21 pagesAlimentos TransgenicosLuis Eduardo Sabastizagal Huerta100% (1)
- Recetario La Mejor Receta Tumbes 2017Document59 pagesRecetario La Mejor Receta Tumbes 2017fiore_l1v8Pas encore d'évaluation
- Triptico de Herbalife PregoneroDocument2 pagesTriptico de Herbalife PregoneroViridianaMendoza100% (1)
- Sociedad Interamericana de Cardiologia - Ecocardio - TEP 2019Document8 pagesSociedad Interamericana de Cardiologia - Ecocardio - TEP 2019Guillermo Mancho CebrianPas encore d'évaluation
- ERAUSQUIN Modelos de Intervención Psicoeducativa Sobre Modelos, Estrategias y Modalidades de Intervención PsicoeducativaDocument23 pagesERAUSQUIN Modelos de Intervención Psicoeducativa Sobre Modelos, Estrategias y Modalidades de Intervención PsicoeducativaClaudia GomezPas encore d'évaluation
- Reglamento Internos ShdugDocument24 pagesReglamento Internos ShdugWillian Ocaña100% (2)
- Aspectos Fundamentales CitocromoDocument177 pagesAspectos Fundamentales CitocromoOctavio Carbajal100% (1)
- Cetoacidosis DiabéticaDocument55 pagesCetoacidosis DiabéticaKimi Jauregui SorianoPas encore d'évaluation
- Restos y OdontogenesisDocument3 pagesRestos y OdontogenesisPaulo EscalantePas encore d'évaluation
- Escala MDIDocument5 pagesEscala MDISilvia González Céspedes100% (3)