Vous aimerez peut-être aussi
- Role of Molecular Chaperones on Structural Folding, Biological Functions, and Drug Interactions of Client ProteinsD'EverandRole of Molecular Chaperones on Structural Folding, Biological Functions, and Drug Interactions of Client ProteinsÉvaluation : 2 sur 5 étoiles2/5 (1)
- Review Article Immunotherapy in The Treatment of Metastatic Melanoma: Current Knowledge and Future DirectionsDocument16 pagesReview Article Immunotherapy in The Treatment of Metastatic Melanoma: Current Knowledge and Future DirectionsSeprianto RuslanPas encore d'évaluation
- Apoptosis and CaspasesDocument19 pagesApoptosis and CaspasesKunhutty Vasudevan AnubhamaPas encore d'évaluation
- The Biology of Apoptosis: Fouad Boulos, MD August 2010Document4 pagesThe Biology of Apoptosis: Fouad Boulos, MD August 2010ans11Pas encore d'évaluation
- Review Mechanisms of Caspase Activation and Inhibition During ApoptosisDocument12 pagesReview Mechanisms of Caspase Activation and Inhibition During ApoptosisFery WirawanPas encore d'évaluation
- ABT-199, A Potent and Selective BCL-2 Inhibitor, Achieves Antitumor Activity While Sparing PlateletsDocument9 pagesABT-199, A Potent and Selective BCL-2 Inhibitor, Achieves Antitumor Activity While Sparing PlateletsSanziana CristeaPas encore d'évaluation
- Principles of Diuretic Therapy: Dr. Rania Magadmi, MBBS, PHDDocument24 pagesPrinciples of Diuretic Therapy: Dr. Rania Magadmi, MBBS, PHDمشاعرمبعثرةPas encore d'évaluation
- Drugs in LactationDocument36 pagesDrugs in Lactationcorzpun168678100% (1)
- Antithyroid Drugs by Sejal Khuman Advanced phARMACOLGY 2Document29 pagesAntithyroid Drugs by Sejal Khuman Advanced phARMACOLGY 2Sejal khumanPas encore d'évaluation
- Eukaryotic DNA ReplicationDocument25 pagesEukaryotic DNA ReplicationKimberly Thomas100% (1)
- Chromatin RemodelingDocument5 pagesChromatin RemodelingRohit GargPas encore d'évaluation
- Renal PharmacokineticsDocument20 pagesRenal PharmacokineticsChristopher VũPas encore d'évaluation
- Carbohydrates MetabolismDocument6 pagesCarbohydrates MetabolismAisha ShahfiqueePas encore d'évaluation
- Genanat D&R AgamDocument51 pagesGenanat D&R AgamWitty GreenPas encore d'évaluation
- Protein Folding - IDocument3 pagesProtein Folding - IJENIFER PEARLIN0% (1)
- General Translation MechanismDocument15 pagesGeneral Translation MechanismAishwarya KashyapPas encore d'évaluation
- Rahimov y Kunkel, 2013Document12 pagesRahimov y Kunkel, 2013Astrith Hernandez Mendoza100% (1)
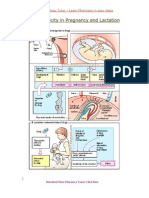
- Drug Toxicity in Pregnancy and LactationDocument3 pagesDrug Toxicity in Pregnancy and LactationPardeep SonyPas encore d'évaluation
- Metabolic Biochemistry, Volume - T. P. MommsenDocument511 pagesMetabolic Biochemistry, Volume - T. P. MommsenJesus Daniel Morales MarquezPas encore d'évaluation
- Enzymatic Test Reaction Biochem Report Group 12Document7 pagesEnzymatic Test Reaction Biochem Report Group 12almira garciaPas encore d'évaluation
- Cell Signaling in Space and Time:: Where Proteins Come Together and When They're ApartDocument20 pagesCell Signaling in Space and Time:: Where Proteins Come Together and When They're ApartHumbertoPas encore d'évaluation
- 3 - Physiology and Anatomy of ReproductionDocument13 pages3 - Physiology and Anatomy of ReproductionjuhPas encore d'évaluation
- Premature Ovarian Failure Incidence Risk Factors ADocument4 pagesPremature Ovarian Failure Incidence Risk Factors Aamit danielPas encore d'évaluation
- Biotechnology (BT)Document11 pagesBiotechnology (BT)Deepakshi SahuPas encore d'évaluation
- Plant Biochemistry Module FinalDocument26 pagesPlant Biochemistry Module FinalHina RaufPas encore d'évaluation
- Biotechnological Growth and ApplicationDocument26 pagesBiotechnological Growth and ApplicationMAITREYEE BANERJEEPas encore d'évaluation
- Bcl-2 Family ProteinsDocument25 pagesBcl-2 Family Proteinsማላያላም ማላያላም100% (1)
- EVS033 Environmental Microbiology - Toxilogy PDFDocument285 pagesEVS033 Environmental Microbiology - Toxilogy PDFRahulPas encore d'évaluation
- Assignment: Department: UimltDocument10 pagesAssignment: Department: UimltEm NomanPas encore d'évaluation
- BiophysicsMolecularTechniques PDFDocument116 pagesBiophysicsMolecularTechniques PDFAnwesha BosePas encore d'évaluation
- DBT JRF Exam Paper + Keys PDFDocument51 pagesDBT JRF Exam Paper + Keys PDFPranjali PriyaPas encore d'évaluation
- Physiology of Skin ReceptorsDocument16 pagesPhysiology of Skin ReceptorsJaynier Jayson JayaPas encore d'évaluation
- The OperonDocument54 pagesThe OperonMoses DesaiPas encore d'évaluation
- Syllabus of Bio-Informatics, PUCCDocument14 pagesSyllabus of Bio-Informatics, PUCCMd MohiuddinPas encore d'évaluation
- The Autonomic Nervous System and Its Central ControlDocument15 pagesThe Autonomic Nervous System and Its Central Controlpuchio100% (1)
- Bioinformatics BS MS: Curriculum OFDocument56 pagesBioinformatics BS MS: Curriculum OFSajid Iqbal LibraryPas encore d'évaluation
- Cell JunctionsDocument29 pagesCell JunctionsArjun DahalPas encore d'évaluation
- Translation and Proteins: The Products of TranscriptionDocument10 pagesTranslation and Proteins: The Products of TranscriptionRenz L. SalumbrePas encore d'évaluation
- Cell Adhesion MoleculesDocument14 pagesCell Adhesion MoleculesSecret Agent100% (1)
- Budiman, PROTEIN ENGINEERING, Budiman, Biomedik, 2014Document33 pagesBudiman, PROTEIN ENGINEERING, Budiman, Biomedik, 2014PMIB Matrikulasi FKUI 2018/2019Pas encore d'évaluation
- Biochemistry of AgingDocument8 pagesBiochemistry of AgingManila MedPas encore d'évaluation
- Drug Use During Pregnancy and LactationDocument45 pagesDrug Use During Pregnancy and LactationSayli GorePas encore d'évaluation
- Gene SilencingDocument17 pagesGene SilencingAmbika saloni100% (1)
- CSIR NET Life Sciences Previous Year Solution June 2022 S1 Part BDocument56 pagesCSIR NET Life Sciences Previous Year Solution June 2022 S1 Part Bpiyush raghavPas encore d'évaluation
- 16383Document5 pages16383Lokesh0% (1)
- Lecture 1 - Enzyme & KineticsDocument33 pagesLecture 1 - Enzyme & KineticsZeny NaranjoPas encore d'évaluation
- Lecture 1Document70 pagesLecture 1Niloy GhoshPas encore d'évaluation
- Drug Transport Across Cell Membrane: Dr. Salman H. RizviDocument67 pagesDrug Transport Across Cell Membrane: Dr. Salman H. RizviIrum RafeeqPas encore d'évaluation
- Prepared By: Shella Mae N. Mainit MSF-1Document44 pagesPrepared By: Shella Mae N. Mainit MSF-1Chriseil arts tancioPas encore d'évaluation
- Protein Immunoblotting: (Western Blotting)Document19 pagesProtein Immunoblotting: (Western Blotting)NANDA AHSANIPas encore d'évaluation
- Examination: (Original Question Paper With Answer Key)Document14 pagesExamination: (Original Question Paper With Answer Key)nothing herePas encore d'évaluation
- Preclass Quiz 8 - Fa19 - MOLECULAR BIOLOGY (47940) PDFDocument3 pagesPreclass Quiz 8 - Fa19 - MOLECULAR BIOLOGY (47940) PDFElizabeth DouglasPas encore d'évaluation
- Pluripotent Stem Cell GuidebookDocument52 pagesPluripotent Stem Cell GuidebookJonathan Torralba TorrónPas encore d'évaluation
- RNA ReplicationDocument23 pagesRNA ReplicationDharaneeshwari Siva-F&NPas encore d'évaluation
- What Is Genome ?Document9 pagesWhat Is Genome ?Bhumi Gameti100% (1)
- LONGLIVE Lifestyle To Conquer AgingDocument21 pagesLONGLIVE Lifestyle To Conquer AgingRakesh SharmaPas encore d'évaluation
- Human Genetics Exam QuestionsDocument10 pagesHuman Genetics Exam Questionsreymond131288Pas encore d'évaluation
- Micro Microbial Mechanisms of Pathogenicity NOTESDocument6 pagesMicro Microbial Mechanisms of Pathogenicity NOTESAriane NoblePas encore d'évaluation
- Lecture 1 IntroductionDocument44 pagesLecture 1 IntroductionAlex HebertPas encore d'évaluation
- The Cell 6Document12 pagesThe Cell 6ZagorkaStambolicPas encore d'évaluation
- Mitochondrial Inheritance (Maternal Inheritance)Document36 pagesMitochondrial Inheritance (Maternal Inheritance)Babak NamiPas encore d'évaluation
- Inheritance of Autosomal Dominant and XlinkedDocument43 pagesInheritance of Autosomal Dominant and XlinkedBabak NamiPas encore d'évaluation
- MosaicismDocument25 pagesMosaicismBabak NamiPas encore d'évaluation
- HemoglobinopathiesDocument37 pagesHemoglobinopathiesBabak Nami100% (1)
- Chromosome Nactivation: Babak Nami Department of Medical Genetics Selçuk UniversityDocument19 pagesChromosome Nactivation: Babak Nami Department of Medical Genetics Selçuk UniversityBabak Nami100% (1)
- Nucleic AcidsDocument113 pagesNucleic AcidsBabak Nami100% (3)
- Uniparental DisomyDocument17 pagesUniparental DisomyBabak NamiPas encore d'évaluation
- Exosomes For Medicine: Babak NamiDocument50 pagesExosomes For Medicine: Babak NamiBabak NamiPas encore d'évaluation
- Human Chromosome NomenclatureDocument35 pagesHuman Chromosome NomenclatureBabak Nami91% (22)
- Aplications of Colchicine As An Article in Medicine: by Babak NamiDocument29 pagesAplications of Colchicine As An Article in Medicine: by Babak NamiBabak NamiPas encore d'évaluation
- Molecular Techniques in Human GeneticsDocument31 pagesMolecular Techniques in Human GeneticsBabak NamiPas encore d'évaluation
- X Inactivation Babak NamiDocument19 pagesX Inactivation Babak NamiBabak NamiPas encore d'évaluation
- GenomeEditingTechniques TheToolsThatEnableScientiststoAltertheGeneticCode EditedDocument14 pagesGenomeEditingTechniques TheToolsThatEnableScientiststoAltertheGeneticCode Editednoneivy anonymousPas encore d'évaluation
- Grading Rubric For DNA ModelDocument2 pagesGrading Rubric For DNA ModelDebbie Ann LaguindabPas encore d'évaluation
- Fpni PCRDocument22 pagesFpni PCRChristianFaltadoCantosPas encore d'évaluation
- Forensics EvidenceDocument80 pagesForensics EvidenceSony BaniyaPas encore d'évaluation
- Chromosomes and DnaDocument5 pagesChromosomes and DnaS. AnsariPas encore d'évaluation
- Compressed Notes Chapter 6: Expression of Biological Information Sb015Document10 pagesCompressed Notes Chapter 6: Expression of Biological Information Sb015SYAZWAN BIN MUSTAFA MoePas encore d'évaluation
- Quarter 3 - MELC 2 Week 2: Biotechnology Activity SheetDocument7 pagesQuarter 3 - MELC 2 Week 2: Biotechnology Activity SheetHeena Grace Presidente100% (1)
- Topic: Base Pair: Hydrogen Bonding and StabilityDocument5 pagesTopic: Base Pair: Hydrogen Bonding and StabilityVINDHYA SHANKERPas encore d'évaluation
- Dream Taq Green MM #k1081Document2 pagesDream Taq Green MM #k1081victor_david_19Pas encore d'évaluation
- Lac Operon PPT FinalDocument18 pagesLac Operon PPT Finalkratikaagrawal50Pas encore d'évaluation
- Hole's Essentials of Human Anatomy & Physiology Twelfth Edition - Chapter 4 Lecture OutlineDocument47 pagesHole's Essentials of Human Anatomy & Physiology Twelfth Edition - Chapter 4 Lecture OutlineCharles AvilaPas encore d'évaluation
- HIV-1 Reverse TranscriptaseDocument3 pagesHIV-1 Reverse TranscriptaseJoshua ArrojoPas encore d'évaluation
- History of The Discovery of DnaDocument15 pagesHistory of The Discovery of DnaEko SujatmikoPas encore d'évaluation
- Functions of OriDocument8 pagesFunctions of OriFelix SimsungPas encore d'évaluation
- Class 12 - Genetics NotesDocument1 pageClass 12 - Genetics NotesDimas HernadyPas encore d'évaluation
- Chapter 8 Study GuideDocument22 pagesChapter 8 Study Guidegabisantana2400Pas encore d'évaluation
- Microbiology A Systems Approach 4Th Edition Cowan Test Bank Full Chapter PDFDocument55 pagesMicrobiology A Systems Approach 4Th Edition Cowan Test Bank Full Chapter PDFChristinaBurketrfd100% (10)
- 8.5 Power Notes PDFDocument1 page8.5 Power Notes PDFMaria Jose SantamariaPas encore d'évaluation
- PCR, RT-PCR, Nested-Pcr, Multiplex PCR, Quantitative PCRDocument116 pagesPCR, RT-PCR, Nested-Pcr, Multiplex PCR, Quantitative PCRYunizardiPas encore d'évaluation
- DNA TranslastionDocument2 pagesDNA TranslastionJoySe Merrera ÜPas encore d'évaluation
- Characterization of Complete Mitochondrial Genomes of Indigenous Mayans in Mexico - ContentServer - AspDocument8 pagesCharacterization of Complete Mitochondrial Genomes of Indigenous Mayans in Mexico - ContentServer - Aspcherrylifev1Pas encore d'évaluation
- Drosophila Genotyping Using PCR: Medya Badakhshani Nikola IlievDocument7 pagesDrosophila Genotyping Using PCR: Medya Badakhshani Nikola IlievSyed Hammad ArfiPas encore d'évaluation
- Dna FingerprintingDocument17 pagesDna Fingerprintingzainab.syed017Pas encore d'évaluation
- 16 DNA StructureDocument27 pages16 DNA StructureAubreyPas encore d'évaluation
- Mechanisms of Long Noncoding RNA Function in Development and DiseaseDocument19 pagesMechanisms of Long Noncoding RNA Function in Development and DiseaseManuel HernándezPas encore d'évaluation
- Lesson 1.7 Learning Task WorksheetDocument6 pagesLesson 1.7 Learning Task WorksheetRayan abdallaPas encore d'évaluation
- Unit 4 Nucleic Acid and Miscellaneous Drug TargetsDocument7 pagesUnit 4 Nucleic Acid and Miscellaneous Drug TargetsCarmela Nadine Grace CuaresmaPas encore d'évaluation
- DV200 Technote Truseq Rna AccessDocument4 pagesDV200 Technote Truseq Rna AccessIoannis PanagopoulosPas encore d'évaluation
- General PCR ProtocolDocument2 pagesGeneral PCR ProtocolSanju KaramPas encore d'évaluation
- Basic Science in Obstetrics and Gynaecology Kak ViraDocument213 pagesBasic Science in Obstetrics and Gynaecology Kak Virakasimsem7Pas encore d'évaluation
- It's Elemental: The Hidden Chemistry in EverythingD'EverandIt's Elemental: The Hidden Chemistry in EverythingÉvaluation : 4 sur 5 étoiles4/5 (10)
- The Nature of Drugs Vol. 1: History, Pharmacology, and Social ImpactD'EverandThe Nature of Drugs Vol. 1: History, Pharmacology, and Social ImpactÉvaluation : 5 sur 5 étoiles5/5 (5)
- Periodic Tales: A Cultural History of the Elements, from Arsenic to ZincD'EverandPeriodic Tales: A Cultural History of the Elements, from Arsenic to ZincÉvaluation : 3.5 sur 5 étoiles3.5/5 (137)
- AP® Chemistry Crash Course, For the 2020 Exam, Book + Online: Get a Higher Score in Less TimeD'EverandAP® Chemistry Crash Course, For the 2020 Exam, Book + Online: Get a Higher Score in Less TimeÉvaluation : 5 sur 5 étoiles5/5 (1)
- Chemistry for Breakfast: The Amazing Science of Everyday LifeD'EverandChemistry for Breakfast: The Amazing Science of Everyday LifeÉvaluation : 4.5 sur 5 étoiles4.5/5 (14)
- Taste: Surprising Stories and Science About Why Food Tastes GoodD'EverandTaste: Surprising Stories and Science About Why Food Tastes GoodÉvaluation : 3 sur 5 étoiles3/5 (20)
- The Periodic Table: A Very Short IntroductionD'EverandThe Periodic Table: A Very Short IntroductionÉvaluation : 4.5 sur 5 étoiles4.5/5 (3)
- Guidelines for Defining Process Safety Competency RequirementsD'EverandGuidelines for Defining Process Safety Competency RequirementsÉvaluation : 3 sur 5 étoiles3/5 (1)
- Is That a Fact?: Frauds, Quacks, and the Real Science of Everyday LifeD'EverandIs That a Fact?: Frauds, Quacks, and the Real Science of Everyday LifeÉvaluation : 5 sur 5 étoiles5/5 (4)
- The Disappearing Spoon: And Other True Tales of Madness, Love, and the History of the World from the Periodic Table of the ElementsD'EverandThe Disappearing Spoon: And Other True Tales of Madness, Love, and the History of the World from the Periodic Table of the ElementsÉvaluation : 4 sur 5 étoiles4/5 (146)
- Monkeys, Myths, and Molecules: Separating Fact from Fiction, and the Science of Everyday LifeD'EverandMonkeys, Myths, and Molecules: Separating Fact from Fiction, and the Science of Everyday LifeÉvaluation : 4 sur 5 étoiles4/5 (1)
- Chemistry for Breakfast: The Amazing Science of Everyday LifeD'EverandChemistry for Breakfast: The Amazing Science of Everyday LifeÉvaluation : 4.5 sur 5 étoiles4.5/5 (90)
- Ingredients: A Visual Exploration of 75 Additives & 25 Food ProductsD'EverandIngredients: A Visual Exploration of 75 Additives & 25 Food ProductsÉvaluation : 4 sur 5 étoiles4/5 (1)
- AP Chemistry Flashcards, Fourth Edition: Up-to-Date Review and PracticeD'EverandAP Chemistry Flashcards, Fourth Edition: Up-to-Date Review and PracticePas encore d'évaluation
- The Nature of Drugs Vol. 1: History, Pharmacology, and Social ImpactD'EverandThe Nature of Drugs Vol. 1: History, Pharmacology, and Social ImpactÉvaluation : 5 sur 5 étoiles5/5 (1)
- Handbook of Formulating Dermal Applications: A Definitive Practical GuideD'EverandHandbook of Formulating Dermal Applications: A Definitive Practical GuidePas encore d'évaluation
- The Regenerative Grower's Guide to Garden Amendments: Using Locally Sourced Materials to Make Mineral and Biological Extracts and FermentsD'EverandThe Regenerative Grower's Guide to Garden Amendments: Using Locally Sourced Materials to Make Mineral and Biological Extracts and FermentsÉvaluation : 5 sur 5 étoiles5/5 (3)
- Chemistry: a QuickStudy Laminated Reference GuideD'EverandChemistry: a QuickStudy Laminated Reference GuideÉvaluation : 5 sur 5 étoiles5/5 (1)
- Tribology: Friction and Wear of Engineering MaterialsD'EverandTribology: Friction and Wear of Engineering MaterialsÉvaluation : 5 sur 5 étoiles5/5 (1)
- Fundamentals of Chemistry: A Modern IntroductionD'EverandFundamentals of Chemistry: A Modern IntroductionÉvaluation : 5 sur 5 étoiles5/5 (1)
- Formulating, Packaging, and Marketing of Natural Cosmetic ProductsD'EverandFormulating, Packaging, and Marketing of Natural Cosmetic ProductsPas encore d'évaluation
- Organic Chemistry for Schools: Advanced Level and Senior High SchoolD'EverandOrganic Chemistry for Schools: Advanced Level and Senior High SchoolPas encore d'évaluation