Vous aimerez peut-être aussi
- Cours Biostat 1AM 15 Sept 2015Document42 pagesCours Biostat 1AM 15 Sept 2015Adonis Serghini100% (2)
- 03 Stratégies Cognitives - 1AMDocument6 pages03 Stratégies Cognitives - 1AMAdonis Serghini100% (1)
- Initiation Vocabulaire-Médical-2015 FMPRDocument37 pagesInitiation Vocabulaire-Médical-2015 FMPRAdonis SerghiniPas encore d'évaluation
- Stat Descriptive1Document25 pagesStat Descriptive1Adonis SerghiniPas encore d'évaluation
- 01 Courants Théoriques - Syllabus 1AMDocument6 pages01 Courants Théoriques - Syllabus 1AMAdonis SerghiniPas encore d'évaluation
- TP Ostéo 2015-2016 EL GHACHI Version AlphaDocument29 pagesTP Ostéo 2015-2016 EL GHACHI Version AlphaAdonis Serghini67% (3)
- 00 Module Methodes Apprentissage - 1AMDocument4 pages00 Module Methodes Apprentissage - 1AMAdonis SerghiniPas encore d'évaluation
- 5 Articulation Scapulo HumeraleDocument3 pages5 Articulation Scapulo HumeraleAdonis SerghiniPas encore d'évaluation
- Résumé D - Ostéologie S1Document20 pagesRésumé D - Ostéologie S1Adonis Serghini100% (2)
- Slides Prévues Pour La Première Séance Biochimie StructuraleDocument34 pagesSlides Prévues Pour La Première Séance Biochimie StructuraleAdonis Serghini100% (1)
- Objectifs 1 Ère Année MédecineDocument60 pagesObjectifs 1 Ère Année MédecineAdonis Serghini86% (7)
- 7-L'Articulation Du PoignetDocument2 pages7-L'Articulation Du PoignetAdonis SerghiniPas encore d'évaluation
- TSG 1Document125 pagesTSG 1Adonis SerghiniPas encore d'évaluation
- URETEREDocument26 pagesURETEREAdonis Serghini88% (8)
- Les CôtesDocument2 pagesLes CôtesAdonis SerghiniPas encore d'évaluation
- Omoplate (Scapula)Document2 pagesOmoplate (Scapula)Adonis SerghiniPas encore d'évaluation
- La ClaviculeDocument2 pagesLa ClaviculeAdonis SerghiniPas encore d'évaluation
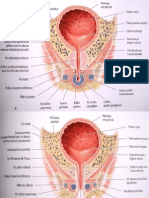
- Organes GénitauxDocument66 pagesOrganes GénitauxAdonis Serghini100% (1)
- Résumé D - Ostéologie S1Document20 pagesRésumé D - Ostéologie S1Adonis Serghini100% (2)
- Tissu Osseux 2Document35 pagesTissu Osseux 2Adonis Serghini100% (2)
- TSG 2.11Document40 pagesTSG 2.11Adonis SerghiniPas encore d'évaluation
- TSG 1Document125 pagesTSG 1Adonis SerghiniPas encore d'évaluation
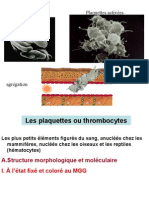
- Tissu SaunguinDocument32 pagesTissu SaunguinAdonis SerghiniPas encore d'évaluation
- Tissu Musculaire 1Document92 pagesTissu Musculaire 1Adonis Serghini100% (2)
- T Nerveux2Document50 pagesT Nerveux2Adonis SerghiniPas encore d'évaluation
- T. Nerveux 3. 1pptDocument50 pagesT. Nerveux 3. 1pptAdonis SerghiniPas encore d'évaluation
- Paroi Thor 2010Document86 pagesParoi Thor 2010Adonis Serghini100% (13)
- Guide Reporting Tableaux de Bord Microsoft Dynamics CRM 2013Document78 pagesGuide Reporting Tableaux de Bord Microsoft Dynamics CRM 2013youssef MCHPas encore d'évaluation
- Notice FH Alcatel 9400uxDocument63 pagesNotice FH Alcatel 9400uxMichelchaPas encore d'évaluation
- Cours 6 Moteurs A Combustion InternesDocument7 pagesCours 6 Moteurs A Combustion Internesahmed ahmadPas encore d'évaluation
- Pages From ASIRI-Recommandations-2012 - CompressedDocument190 pagesPages From ASIRI-Recommandations-2012 - CompressedYaseen AlAjmi CompanyPas encore d'évaluation
- Notes de Cours BD 27-07-2023Document3 pagesNotes de Cours BD 27-07-2023Aimé ZahuiPas encore d'évaluation
- Guide D'administration TWSDocument413 pagesGuide D'administration TWSDenazareth JesusPas encore d'évaluation
- QCM1Document4 pagesQCM1ChristianPas encore d'évaluation
- PL - Chap I - Copie PDFDocument5 pagesPL - Chap I - Copie PDFmme tPas encore d'évaluation
- L Etude de La Geolocalisation Dans Les Reseaux Mobiles GSM Et CDMA A MOBILINFODocument77 pagesL Etude de La Geolocalisation Dans Les Reseaux Mobiles GSM Et CDMA A MOBILINFOAdama SowPas encore d'évaluation
- Cours Theorie Du Controle3Document38 pagesCours Theorie Du Controle3Abderrahim El amraniPas encore d'évaluation
- Memoire de Fin D'EtudesDocument63 pagesMemoire de Fin D'Etudestakoua.belghitPas encore d'évaluation
- RCI BEPC 2016 Zone3 Mathematiques PDFDocument2 pagesRCI BEPC 2016 Zone3 Mathematiques PDFNET MONEYPas encore d'évaluation
- 2022 NelleCaledo SI J2 Exo3 Sujet PAC 10ptsDocument3 pages2022 NelleCaledo SI J2 Exo3 Sujet PAC 10ptsgabinPas encore d'évaluation
- Sujet 03Document4 pagesSujet 03lbog.trashPas encore d'évaluation
- DS1b PCDocument3 pagesDS1b PCfrancoisbPas encore d'évaluation
- Pfe GC 0012-7Document10 pagesPfe GC 0012-7Gifted MouhcinePas encore d'évaluation
- Les VecteursDocument6 pagesLes VecteursGAEL ETIENNEPas encore d'évaluation
- TD Thermo SERIE2 AIT HOUDocument2 pagesTD Thermo SERIE2 AIT HOUOum FirasPas encore d'évaluation
- Radiologie Anul IIIDocument47 pagesRadiologie Anul IIIlolcPas encore d'évaluation
- QCM Corro Architecture - 123325Document6 pagesQCM Corro Architecture - 123325Grâce AudreyPas encore d'évaluation
- Corrigé Type Du Modue Les Écoulements en Charge Master 1 Hydraulique UrbaineDocument5 pagesCorrigé Type Du Modue Les Écoulements en Charge Master 1 Hydraulique UrbaineMimouna TabetPas encore d'évaluation
- Filtration GlomerulaireDocument16 pagesFiltration GlomerulaireRania HamdiPas encore d'évaluation
- Cours Materiaux Isolants - DocDocument35 pagesCours Materiaux Isolants - DocJihen Rf100% (1)
- IntroductionDocument8 pagesIntroductionMezine SohaibPas encore d'évaluation
- 16 17 Examen BD Smi Lpii CorrectionDocument6 pages16 17 Examen BD Smi Lpii Correctionadilcharahil2000Pas encore d'évaluation
- Les Environnements SedimentairesDocument22 pagesLes Environnements SedimentairesKhnati RidaPas encore d'évaluation
- Chap1 TP1 Python en Interactif (1) GRGGDocument6 pagesChap1 TP1 Python en Interactif (1) GRGGMathis .LPas encore d'évaluation
- Examen Blanc 2Document4 pagesExamen Blanc 2AlcalinaPhysicsPas encore d'évaluation
- Analyse Granulométrique Compte Rendu TP MDC Génie Civil PDFDocument10 pagesAnalyse Granulométrique Compte Rendu TP MDC Génie Civil PDFtatif100% (5)
- S61 - ENPU Synthese CVSDocument9 pagesS61 - ENPU Synthese CVSBahija KOUHAILPas encore d'évaluation