Vous aimerez peut-être aussi
- Coloration Simple Alina ClimaDocument3 pagesColoration Simple Alina ClimaAlina VetriceanPas encore d'évaluation
- Dyn PMDocument8 pagesDyn PMAlina VetriceanPas encore d'évaluation
- Travail de Laboratoire (Met-Fiz.) nr.1 ClimaDocument3 pagesTravail de Laboratoire (Met-Fiz.) nr.1 ClimaAlina VetriceanPas encore d'évaluation
- Lucrare de Laborator ClimaDocument6 pagesLucrare de Laborator ClimaAlina VetriceanPas encore d'évaluation
- Calcules 1 ClimaDocument3 pagesCalcules 1 ClimaAlina VetriceanPas encore d'évaluation
- Cin. PMDocument10 pagesCin. PMAlina VetriceanPas encore d'évaluation
- Les OsidesDocument41 pagesLes OsidesAlina VetriceanPas encore d'évaluation
- Museum D Histoire NaturelleDocument6 pagesMuseum D Histoire NaturelleAlina VetriceanPas encore d'évaluation
- BordeauxDocument7 pagesBordeauxAlina VetriceanPas encore d'évaluation
- L'économie Des Pays FrancophonesDocument43 pagesL'économie Des Pays FrancophonesAlina VetriceanPas encore d'évaluation
- PatiserieDocument18 pagesPatiserieAlina VetriceanPas encore d'évaluation
- Salmonella (Text)Document4 pagesSalmonella (Text)Alina VetriceanPas encore d'évaluation
- Les Deux MagotsDocument9 pagesLes Deux MagotsAlina VetriceanPas encore d'évaluation
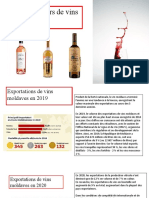
- Exportations de Vins MoldavesDocument4 pagesExportations de Vins MoldavesAlina VetriceanPas encore d'évaluation
- AminoaciziiDocument3 pagesAminoaciziiAlina VetriceanPas encore d'évaluation
- 2.4 Réactions Chimiques Des GlucidesDocument2 pages2.4 Réactions Chimiques Des GlucidesAlina VetriceanPas encore d'évaluation
- Controlul Microbiologic Al CărniiDocument9 pagesControlul Microbiologic Al CărniiAlina VetriceanPas encore d'évaluation
- Calitatea Nutrițională A ProteinelorDocument2 pagesCalitatea Nutrițională A ProteinelorAlina VetriceanPas encore d'évaluation
- Exemple de ProteineDocument3 pagesExemple de ProteineAlina VetriceanPas encore d'évaluation
- 2.3 Osides. SaccharidesDocument8 pages2.3 Osides. SaccharidesAlina VetriceanPas encore d'évaluation
- VitaminesDocument19 pagesVitaminesAlina VetriceanPas encore d'évaluation
- Chimie Alimentară 3Document2 pagesChimie Alimentară 3Alina VetriceanPas encore d'évaluation
- Microbiologie (Atest 2)Document9 pagesMicrobiologie (Atest 2)Alina VetriceanPas encore d'évaluation
- Degradarea ProteinelorDocument2 pagesDegradarea ProteinelorAlina VetriceanPas encore d'évaluation
- PeptideleDocument2 pagesPeptideleAlina VetriceanPas encore d'évaluation
- Politique de ProduitDocument39 pagesPolitique de ProduitAlina VetriceanPas encore d'évaluation
- Les LipidesDocument20 pagesLes LipidesAlina VetriceanPas encore d'évaluation
- Le MarketingDocument25 pagesLe MarketingAlina VetriceanPas encore d'évaluation
- Voie Des Pentoses Phosphates (MaJ) - 2Document1 pageVoie Des Pentoses Phosphates (MaJ) - 2Hadrien De Greef100% (2)
- Ferrite 16 PDFDocument3 pagesFerrite 16 PDFanoosh nymousPas encore d'évaluation
- Contrôle 2, TCS BIOF, 2017 - 2018Document3 pagesContrôle 2, TCS BIOF, 2017 - 2018khalifa El harfiPas encore d'évaluation
- SME CaoutchoucDocument113 pagesSME CaoutchoucsusCitiesPas encore d'évaluation
- Méthode de Dosage Des Vitamines Hydrosolubles PDFDocument40 pagesMéthode de Dosage Des Vitamines Hydrosolubles PDFHamza AkirPas encore d'évaluation
- Les ProteinesDocument10 pagesLes ProteinesWissal ElbarPas encore d'évaluation
- Exercice PCRDocument1 pageExercice PCRSalima saidPas encore d'évaluation
- Lait 54 1974 535-536 11Document9 pagesLait 54 1974 535-536 11yassine tijaniPas encore d'évaluation
- Fle040 Geranium Bourbon IfraDocument4 pagesFle040 Geranium Bourbon IfraGilPas encore d'évaluation
- Fichier Produit 2954Document46 pagesFichier Produit 2954Chokri PharmPas encore d'évaluation
- Doctorat en Pectine en Pharmacie FDocument111 pagesDoctorat en Pectine en Pharmacie FHAMZA AZOUAITINPas encore d'évaluation
- INTRODUCTIONDocument6 pagesINTRODUCTIONYvan Cedric AgoPas encore d'évaluation
- Janv 2017Document4 pagesJanv 2017ashPas encore d'évaluation
- Polycopié de TP Enzymologie - PrHammani - M21 SVI4 2021 2022Document12 pagesPolycopié de TP Enzymologie - PrHammani - M21 SVI4 2021 2022mohamed KhnifratPas encore d'évaluation
- Viscosity of Inorganic and Organic LiquidsDocument7 pagesViscosity of Inorganic and Organic LiquidsGabriel SugayaPas encore d'évaluation
- Cours de MetabolismeDocument69 pagesCours de Metabolismeleonce kadjiPas encore d'évaluation
- C5 - Structure Et Caractérisation Des Molécules OrganiquesDocument4 pagesC5 - Structure Et Caractérisation Des Molécules OrganiquesloukaPas encore d'évaluation
- Chap IIDocument12 pagesChap IIfadoua lakouissiPas encore d'évaluation
- TP S4 Nitrate Réductase Printemps 2022Document19 pagesTP S4 Nitrate Réductase Printemps 2022khaoula.banouissPas encore d'évaluation
- Poly GIM 2021-2022 E. Richaud Cours 2 + 3 Mars 2022 - FelipeDocument20 pagesPoly GIM 2021-2022 E. Richaud Cours 2 + 3 Mars 2022 - FelipeMateus Pio CarneiroPas encore d'évaluation
- Protocole de preparation-GHADocument5 pagesProtocole de preparation-GHAJean Paul Ndongo OkalaPas encore d'évaluation
- Food SI 2023 GK Practice Set 003 by Tapas KumbhakarDocument4 pagesFood SI 2023 GK Practice Set 003 by Tapas Kumbhakarkaustav royPas encore d'évaluation
- TP 1 Détermination de L'amidon Par Méthode PolarimétriqueDocument7 pagesTP 1 Détermination de L'amidon Par Méthode PolarimétriqueAmirou Baby Mixico100% (4)
- Devoir de Contrôle N°1 2017 2018 (MR Salhi Mohamed)Document2 pagesDevoir de Contrôle N°1 2017 2018 (MR Salhi Mohamed)Wiem HabbassiPas encore d'évaluation
- CoursDocument22 pagesCourspouambeka tony wheellyamPas encore d'évaluation
- Correction Exercices Spectres UV Visible IRDocument3 pagesCorrection Exercices Spectres UV Visible IRImad MadaniPas encore d'évaluation
- HeteroDocument35 pagesHeteroergrt hdfhdhPas encore d'évaluation
- Ecologie G 4Document11 pagesEcologie G 4Samy IbkPas encore d'évaluation
- PESTICIDESDocument21 pagesPESTICIDESpc ghomriPas encore d'évaluation
- L'analyse Du Vin: A - Détermination Du Degré Alcoolique Du VinDocument15 pagesL'analyse Du Vin: A - Détermination Du Degré Alcoolique Du Vinmagloire amivaPas encore d'évaluation
- Harmonisation Energétique des Lieux: Habitat et haut-lieux sacrés 2020D'EverandHarmonisation Energétique des Lieux: Habitat et haut-lieux sacrés 2020Évaluation : 2.5 sur 5 étoiles2.5/5 (3)
- Améliorer votre mémoire: Un Guide pour l'augmentation de la puissance du cerveau, utilisant des techniques et méthodesD'EverandAméliorer votre mémoire: Un Guide pour l'augmentation de la puissance du cerveau, utilisant des techniques et méthodesÉvaluation : 5 sur 5 étoiles5/5 (2)
- La vie des abeilles: Prix Nobel de littératureD'EverandLa vie des abeilles: Prix Nobel de littératureÉvaluation : 4 sur 5 étoiles4/5 (41)
- Technologie automobile: Les Grands Articles d'UniversalisD'EverandTechnologie automobile: Les Grands Articles d'UniversalisPas encore d'évaluation
- Manuel pour les débutants Fabriquez des savons naturelsD'EverandManuel pour les débutants Fabriquez des savons naturelsÉvaluation : 3 sur 5 étoiles3/5 (2)
- Secrets ancestraux d'un maître guérisseur: Un sceptique occidental, un maître oriental et les plus grands secrets de la vieD'EverandSecrets ancestraux d'un maître guérisseur: Un sceptique occidental, un maître oriental et les plus grands secrets de la vieÉvaluation : 5 sur 5 étoiles5/5 (2)
- Transformez votre vie: Utilisez le pouvoir créateur qui est en vous pour construire votre vie à l'image de ce que vous voulez qu'elle soitD'EverandTransformez votre vie: Utilisez le pouvoir créateur qui est en vous pour construire votre vie à l'image de ce que vous voulez qu'elle soitÉvaluation : 4 sur 5 étoiles4/5 (14)
- 20 Véritables remèdes de nos grands-mères pour maigrir vite et enfin perdre du poidsD'Everand20 Véritables remèdes de nos grands-mères pour maigrir vite et enfin perdre du poidsÉvaluation : 5 sur 5 étoiles5/5 (1)
- Approvisionnement et traitement de l’eau: Les Grands Articles d'UniversalisD'EverandApprovisionnement et traitement de l’eau: Les Grands Articles d'UniversalisPas encore d'évaluation
- Semer avec succès pour rassembler avec abundance. Jardin organique et synergique: Calcul des meilleurs jours pour l'ensemencement de chaque légumeD'EverandSemer avec succès pour rassembler avec abundance. Jardin organique et synergique: Calcul des meilleurs jours pour l'ensemencement de chaque légumePas encore d'évaluation
- Géobiologie de l'habitat et Géobiologie sacrée: Pour un lieu sainD'EverandGéobiologie de l'habitat et Géobiologie sacrée: Pour un lieu sainÉvaluation : 4.5 sur 5 étoiles4.5/5 (2)
- Conception & Modélisation CAO: Le guide ultime du débutantD'EverandConception & Modélisation CAO: Le guide ultime du débutantPas encore d'évaluation
- L'Art de la guerre: Traité de stratégie en 13 chapitres (texte intégral)D'EverandL'Art de la guerre: Traité de stratégie en 13 chapitres (texte intégral)Évaluation : 4 sur 5 étoiles4/5 (3032)
- Géologie de l'Amérique: Les Grands Articles d'UniversalisD'EverandGéologie de l'Amérique: Les Grands Articles d'UniversalisPas encore d'évaluation
- 500 secrets pour avoir un potager merveilleuxD'Everand500 secrets pour avoir un potager merveilleuxÉvaluation : 2 sur 5 étoiles2/5 (1)
- L'Ombre à l'Univers: La structure des particules élémentaires XIIfD'EverandL'Ombre à l'Univers: La structure des particules élémentaires XIIfPas encore d'évaluation
- Histoire de la psychologie scientifique: De la naissance de la psychologie à la neuropsychologie et aux champs d'application les plus actuelsD'EverandHistoire de la psychologie scientifique: De la naissance de la psychologie à la neuropsychologie et aux champs d'application les plus actuelsPas encore d'évaluation
- Jus de Fruits et de Légumes Crus: 57 recettes faciles et un Guide Pratique Complet pour améliorer votre alimentation .: Santé, Vitalité et Minceur, avec ... ET DURABLEMENTD'EverandJus de Fruits et de Légumes Crus: 57 recettes faciles et un Guide Pratique Complet pour améliorer votre alimentation .: Santé, Vitalité et Minceur, avec ... ET DURABLEMENTPas encore d'évaluation
- 160 ressources pour se lancer dans la vidéo quand on n’y connait rienD'Everand160 ressources pour se lancer dans la vidéo quand on n’y connait rienPas encore d'évaluation