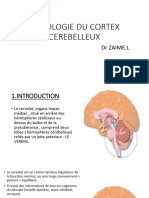
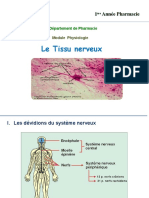
Vous aimerez peut-être aussi
- Neurobiologie: Les Grands Articles d'UniversalisD'EverandNeurobiologie: Les Grands Articles d'UniversalisPas encore d'évaluation
- CM4 - Vascularisation Et Voies Sensitivo-MotricesDocument14 pagesCM4 - Vascularisation Et Voies Sensitivo-Motricescharbelantoun2443Pas encore d'évaluation
- Le Système NerveuxDocument8 pagesLe Système NerveuxazizPas encore d'évaluation
- Le Tissu NerveuxDocument10 pagesLe Tissu NerveuxANDRIANOME tafitaPas encore d'évaluation
- Cours Physiologie Animale 1-1Document150 pagesCours Physiologie Animale 1-1Mina OussarPas encore d'évaluation
- Neurulation Et Modelage Du MésodermeDocument52 pagesNeurulation Et Modelage Du MésodermeGeorge BorsPas encore d'évaluation
- La Quatrième Semaine de DéveloppementDocument54 pagesLa Quatrième Semaine de DéveloppementYacouba DialloPas encore d'évaluation
- La Neurophysiologie Cours 2013Document17 pagesLa Neurophysiologie Cours 2013friends diaryPas encore d'évaluation
- Neurone PDFDocument18 pagesNeurone PDFyasminePas encore d'évaluation
- 4ème Semaine D.EDocument5 pages4ème Semaine D.ENouhoum TraoréPas encore d'évaluation
- Le Cortex Cérébelleux Partie 2Document9 pagesLe Cortex Cérébelleux Partie 2louanne kloepferPas encore d'évaluation
- 1-Résumé Lydia Tadjadit Part1 PDFDocument41 pages1-Résumé Lydia Tadjadit Part1 PDFSaibiPas encore d'évaluation
- 03-Le CerveletDocument5 pages03-Le Cervelethocine benganaPas encore d'évaluation
- Cours SVT Tle D&C 2013-2014Document17 pagesCours SVT Tle D&C 2013-2014sandwidibernard76Pas encore d'évaluation
- Neuroanat-2 0Document83 pagesNeuroanat-2 0seloua kaid100% (1)
- MS 1990 1 64Document2 pagesMS 1990 1 64Fériel LchPas encore d'évaluation
- Cours SN DR NaoualiDocument75 pagesCours SN DR NaoualiKobePas encore d'évaluation
- 4eme Type Tissu NerveuxDocument36 pages4eme Type Tissu Nerveuxgroupe_svi3Pas encore d'évaluation
- 07-Le Tissu NerveuxDocument11 pages07-Le Tissu NerveuxBemmoussat AsmaPas encore d'évaluation
- Generalites IDocument11 pagesGeneralites IzqwjjbngxnPas encore d'évaluation
- 1a d1 Ue8 Cours 1 Developpement Du Systeme Nerveux Central-RoneoDocument14 pages1a d1 Ue8 Cours 1 Developpement Du Systeme Nerveux Central-RoneosyurmiPas encore d'évaluation
- Le CreveletDocument6 pagesLe CreveletŽĭņëb BėñPas encore d'évaluation
- 4 - Histo Du Cortex CérébelleuxDocument21 pages4 - Histo Du Cortex Cérébelleuxbf ImprimeriePas encore d'évaluation
- Physiologie Du NeuroneDocument21 pagesPhysiologie Du NeuronelbawainaPas encore d'évaluation
- Neurophysiologie 2Document49 pagesNeurophysiologie 2123kiwiPas encore d'évaluation
- 1 Er Cour PhysiologieDocument13 pages1 Er Cour Physiologiederdiri imènePas encore d'évaluation
- 3 - Organogenèse, Histiogenèse Et Cytogenèse Du Système Nerveux Central (Cours)Document10 pages3 - Organogenèse, Histiogenèse Et Cytogenèse Du Système Nerveux Central (Cours)YanisDessaillyPas encore d'évaluation
- Le Tissu NerveuxDocument51 pagesLe Tissu NerveuxYouness KhalfaouiPas encore d'évaluation
- Partie 4Document31 pagesPartie 4ndeye yarame diopPas encore d'évaluation
- Ganglion Nerveux FinalDocument3 pagesGanglion Nerveux Finalmalakzerizer2005Pas encore d'évaluation
- Neurobio Mots-Cle s-MargauxTerlindenDocument20 pagesNeurobio Mots-Cle s-MargauxTerlindenDaniela RiosPas encore d'évaluation
- Pocock475819 PDFDocument11 pagesPocock475819 PDFMohamed DaifPas encore d'évaluation
- 01 - Le Tissu NerveuxDocument13 pages01 - Le Tissu NerveuxNesrinePas encore d'évaluation
- Le Tissu Nerveux - Vue GénéraleDocument67 pagesLe Tissu Nerveux - Vue GénéraleSiaka YodaPas encore d'évaluation
- Tissus NerveuxDocument19 pagesTissus NerveuxGuenzet Akila100% (1)
- Neuroanatomie 2Document79 pagesNeuroanatomie 2Tr LwPas encore d'évaluation
- 1 - Mostaganem Polycope Histologie 2012 - 2013 PDFDocument62 pages1 - Mostaganem Polycope Histologie 2012 - 2013 PDFRébecca ElangaPas encore d'évaluation
- 2-Épitheluim GlandulaireDocument13 pages2-Épitheluim GlandulaireDissa SaliaPas encore d'évaluation
- 7 Neurulation Mesoderme 2014 2015 PDFDocument56 pages7 Neurulation Mesoderme 2014 2015 PDFnicolaer111100% (1)
- BIOLE102 Neurobiologie: LégendeDocument117 pagesBIOLE102 Neurobiologie: LégendeDaniela RiosPas encore d'évaluation
- 8-Tissu NerveuxDocument99 pages8-Tissu NerveuxAy ManPas encore d'évaluation
- Uef 102 Diapo Tissu NerveuxDocument123 pagesUef 102 Diapo Tissu NerveuxAlia YmfPas encore d'évaluation
- Physiopathologie NeuroscienceDocument46 pagesPhysiopathologie NeuroscienceYas MiiinaaPas encore d'évaluation
- Biologie Animale P1 Emb NeurulationDocument2 pagesBiologie Animale P1 Emb Neurulationhammou HoudjedjePas encore d'évaluation
- 3-Le Tissu NerveuxDocument19 pages3-Le Tissu Nerveuxboutefal imenePas encore d'évaluation
- Polycopié Histologie Médecine Béchar 1èreDocument30 pagesPolycopié Histologie Médecine Béchar 1èreFaculté De Médecine BécharPas encore d'évaluation
- Le Tissu NerveuxDocument16 pagesLe Tissu NerveuxZizou HamdiPas encore d'évaluation
- Résumé G.Chakib NerveuxDocument5 pagesRésumé G.Chakib NerveuxMaroua AbzPas encore d'évaluation
- 50 41904Document5 pages50 41904salimPas encore d'évaluation
- Planche Otn3 LTSNTDocument2 pagesPlanche Otn3 LTSNTAida Mbodji100% (1)
- La Cellule NerveuseDocument5 pagesLa Cellule Nerveusemohamed1539100% (1)
- L'oeilDocument8 pagesL'oeilmalakzerizer2005Pas encore d'évaluation
- Synthèse La CelluleDocument6 pagesSynthèse La CelluleKamil LahbabiPas encore d'évaluation
- Les ÉpithéliumsDocument2 pagesLes ÉpithéliumsDi BenamoPas encore d'évaluation
- Biologie Cellulaire Partie 1Document29 pagesBiologie Cellulaire Partie 1rhannajezouhair19Pas encore d'évaluation
- Le Noyau Cours SNVDocument9 pagesLe Noyau Cours SNVDear tynaPas encore d'évaluation
- Le Cortex CerebralDocument3 pagesLe Cortex CerebralLolo KawtchiPas encore d'évaluation
- 1.6 - Anatomie Et Physiologie Du Systeme Nerveux.Document24 pages1.6 - Anatomie Et Physiologie Du Systeme Nerveux.Clément IsoardPas encore d'évaluation
- Ms Ebm Benhabib+BradaiDocument63 pagesMs Ebm Benhabib+BradaiDady LalaPas encore d'évaluation
- Biocellsession 2Document15 pagesBiocellsession 2Eroline GuelciPas encore d'évaluation
- Stress Postraumatic 1Document11 pagesStress Postraumatic 1Maria StanicaPas encore d'évaluation
- Saccomplir PleinementDocument76 pagesSaccomplir PleinementAbderrazzak Houdzi100% (2)
- Cours Psychologie CognitiveDocument20 pagesCours Psychologie CognitiveSiham HamidiPas encore d'évaluation
- QCM Adaptabilitecc81 de LorganismeDocument2 pagesQCM Adaptabilitecc81 de Lorganismesohane MdiPas encore d'évaluation
- Le ParfumDocument11 pagesLe ParfumAnonymous nzfEotriWKPas encore d'évaluation
- Troubles NeurodéveloppementauxDocument33 pagesTroubles NeurodéveloppementauxSophie RicherPas encore d'évaluation
- Le Livre de l'Au-Delà - Bô Yin Râ PDFDocument96 pagesLe Livre de l'Au-Delà - Bô Yin Râ PDFMoumouni Ouedraogo100% (2)
- Le Cerveau en 30 Secondes PDFDocument238 pagesLe Cerveau en 30 Secondes PDFLombe Christian100% (1)
- PR Formation Sensorielle Les Cafés DuboisDocument16 pagesPR Formation Sensorielle Les Cafés DuboisAssoum AssoumPas encore d'évaluation
- CM Psycho CognitiveDocument115 pagesCM Psycho CognitiveLaetitia PicardPas encore d'évaluation
- L'examen PsychiatriqueDocument21 pagesL'examen PsychiatriqueAhlam missPas encore d'évaluation
- PNL - Techniques Secretes de Per - Alain PomerleauDocument89 pagesPNL - Techniques Secretes de Per - Alain Pomerleausanamoussa153100% (2)
- Angiopathie Amyloïde CérébraleDocument46 pagesAngiopathie Amyloïde CérébraleIchrak GhachemPas encore d'évaluation
- Deep Learning 1Document20 pagesDeep Learning 1chaimaePas encore d'évaluation
- Diagnostics différentiels des psychoses endogènes, des troubles de la personnalité et des névrosesD'EverandDiagnostics différentiels des psychoses endogènes, des troubles de la personnalité et des névrosesÉvaluation : 4.5 sur 5 étoiles4.5/5 (2)
- Améliorer votre mémoire: Un Guide pour l'augmentation de la puissance du cerveau, utilisant des techniques et méthodesD'EverandAméliorer votre mémoire: Un Guide pour l'augmentation de la puissance du cerveau, utilisant des techniques et méthodesÉvaluation : 5 sur 5 étoiles5/5 (2)
- Guide De Démarrage De L'Apiculture: Le Guide Complet Pour Élever Des AbeillesD'EverandGuide De Démarrage De L'Apiculture: Le Guide Complet Pour Élever Des AbeillesPas encore d'évaluation
- Introduction à la psychologie des émotions: De Darwin aux neurosciences: découvrir les émotions et leur mode de fonctionnementD'EverandIntroduction à la psychologie des émotions: De Darwin aux neurosciences: découvrir les émotions et leur mode de fonctionnementPas encore d'évaluation
- Géologie de l'Amérique: Les Grands Articles d'UniversalisD'EverandGéologie de l'Amérique: Les Grands Articles d'UniversalisPas encore d'évaluation
- La vie des abeilles: Prix Nobel de littératureD'EverandLa vie des abeilles: Prix Nobel de littératureÉvaluation : 4 sur 5 étoiles4/5 (41)
- Harmonisation Energétique des Lieux: Habitat et haut-lieux sacrés 2020D'EverandHarmonisation Energétique des Lieux: Habitat et haut-lieux sacrés 2020Évaluation : 2.5 sur 5 étoiles2.5/5 (3)
- Neuropsychologie: Les bases théoriques et pratiques du domaine d'étude (psychologie pour tous)D'EverandNeuropsychologie: Les bases théoriques et pratiques du domaine d'étude (psychologie pour tous)Pas encore d'évaluation
- Le TDAH chez l’adulte : Comment reconnaître & faire face au TDAH chez l’adulte en 30 étapes faciles.D'EverandLe TDAH chez l’adulte : Comment reconnaître & faire face au TDAH chez l’adulte en 30 étapes faciles.Pas encore d'évaluation
- Le Pouvoir De L'Autodiscipline: L’art de tout accomplir sans limiteD'EverandLe Pouvoir De L'Autodiscipline: L’art de tout accomplir sans limiteÉvaluation : 4 sur 5 étoiles4/5 (16)
- Cahier de jeux de stimulation cognitive: Sujets Alzheimer, désorientés, démences, amnésiesD'EverandCahier de jeux de stimulation cognitive: Sujets Alzheimer, désorientés, démences, amnésiesPas encore d'évaluation
- La thérapie libre: La Thérapie de LIBération et Résilience EmotionnelleD'EverandLa thérapie libre: La Thérapie de LIBération et Résilience EmotionnelleÉvaluation : 5 sur 5 étoiles5/5 (1)
- Hypnose & affirmations positives pour la richesse, l’abondance et la réussite (21 jours pour un nouveau vous)D'EverandHypnose & affirmations positives pour la richesse, l’abondance et la réussite (21 jours pour un nouveau vous)Évaluation : 5 sur 5 étoiles5/5 (1)
- Histoire de la psychologie scientifique: De la naissance de la psychologie à la neuropsychologie et aux champs d'application les plus actuelsD'EverandHistoire de la psychologie scientifique: De la naissance de la psychologie à la neuropsychologie et aux champs d'application les plus actuelsPas encore d'évaluation
- Autisme - La boîte à outils: Stratégies et techniques pour accompagner un enfant autisteD'EverandAutisme - La boîte à outils: Stratégies et techniques pour accompagner un enfant autistePas encore d'évaluation
- Le CODE DE DIEU: Le secret de notre passé, la promesse de notre avenirD'EverandLe CODE DE DIEU: Le secret de notre passé, la promesse de notre avenirÉvaluation : 4 sur 5 étoiles4/5 (23)
- La psychologie de l'anxiété: La connaître pour comprendre ses mécanismes de fonctionnementD'EverandLa psychologie de l'anxiété: La connaître pour comprendre ses mécanismes de fonctionnementPas encore d'évaluation
- Troubles Bipolaires: Mieux les connaître pour mieux se débarrasser de ces souffrancesD'EverandTroubles Bipolaires: Mieux les connaître pour mieux se débarrasser de ces souffrancesÉvaluation : 4.5 sur 5 étoiles4.5/5 (3)
- Qu'arrive t'il a la terre arable ?: Le problème de l'érosion du sol.D'EverandQu'arrive t'il a la terre arable ?: Le problème de l'érosion du sol.Évaluation : 5 sur 5 étoiles5/5 (1)