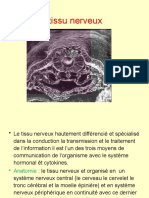
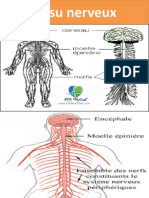
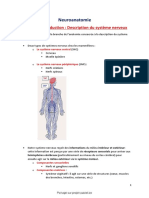
Vous aimerez peut-être aussi
- Le Tissu NerveuxDocument10 pagesLe Tissu NerveuxANDRIANOME tafitaPas encore d'évaluation
- 08 - LE TISSU NERVEUXDocument7 pages08 - LE TISSU NERVEUXlahlahazouPas encore d'évaluation
- 4eme Type Tissu NerveuxDocument36 pages4eme Type Tissu Nerveuxgroupe_svi3Pas encore d'évaluation
- Résumé G.Chakib NerveuxDocument5 pagesRésumé G.Chakib NerveuxMaroua AbzPas encore d'évaluation
- Tissu NerveuxDocument6 pagesTissu Nerveuxtim SssPas encore d'évaluation
- Tissu Nerveux UCADDocument66 pagesTissu Nerveux UCADDiabel AndøPas encore d'évaluation
- Chapitre II: Organisation Cellulaire Du Système NerveuxDocument36 pagesChapitre II: Organisation Cellulaire Du Système Nerveuxnesrine MoadPas encore d'évaluation
- Le Tissu Nerveux 2022Document90 pagesLe Tissu Nerveux 2022babacar98.04Pas encore d'évaluation
- Tissu NerveuxDocument6 pagesTissu NerveuxOussama FendaliPas encore d'évaluation
- Physiologie Du NeuroneDocument21 pagesPhysiologie Du NeuronelbawainaPas encore d'évaluation
- Le Tissu NerveuxDocument16 pagesLe Tissu NerveuxZizou HamdiPas encore d'évaluation
- Cours SVT Tle D&C 2013-2014Document17 pagesCours SVT Tle D&C 2013-2014sandwidibernard76Pas encore d'évaluation
- 1 Er Cour PhysiologieDocument13 pages1 Er Cour Physiologiederdiri imènePas encore d'évaluation
- MI3 Neurosciences Histologie Du Systeme Nerveux LavabreDocument17 pagesMI3 Neurosciences Histologie Du Systeme Nerveux LavabreDjallal HassaniPas encore d'évaluation
- 01 - Le Tissu NerveuxDocument13 pages01 - Le Tissu NerveuxNesrinePas encore d'évaluation
- Le Tissu NerveuxDocument51 pagesLe Tissu NerveuxYouness KhalfaouiPas encore d'évaluation
- Le Tissu Nerveux - Vue GénéraleDocument67 pagesLe Tissu Nerveux - Vue GénéraleSiaka YodaPas encore d'évaluation
- Neurophysiologie 2Document49 pagesNeurophysiologie 2123kiwiPas encore d'évaluation
- La Cellule NerveuseDocument5 pagesLa Cellule Nerveusemohamed1539100% (1)
- Les Tissus NerveuxDocument9 pagesLes Tissus NerveuxZoulkiffiPas encore d'évaluation
- Cours Neurohisto PCEM1 2006-2007Document50 pagesCours Neurohisto PCEM1 2006-2007Wiwi LauraPas encore d'évaluation
- Generalites IDocument11 pagesGeneralites IzqwjjbngxnPas encore d'évaluation
- La Neurophysiologie Cours 2013Document17 pagesLa Neurophysiologie Cours 2013friends diaryPas encore d'évaluation
- Tissu Nerveux - PolycopiéDocument10 pagesTissu Nerveux - PolycopiéAmina DrkPas encore d'évaluation
- Physiologie Du Nerf 2022 Mede UM6Document163 pagesPhysiologie Du Nerf 2022 Mede UM6Girl Ytb100% (1)
- Tissu Nerveux Part1 MelDocument4 pagesTissu Nerveux Part1 MelmelPas encore d'évaluation
- Neuroanatomie Complet JustineDocument123 pagesNeuroanatomie Complet JustineMaria OlteanPas encore d'évaluation
- Cours SN DR NaoualiDocument75 pagesCours SN DR NaoualiKobePas encore d'évaluation
- MOD6 Le Syste Me Nerveux Central - La Moelle e Pinie Re PDFDocument3 pagesMOD6 Le Syste Me Nerveux Central - La Moelle e Pinie Re PDFmedPas encore d'évaluation
- Cours Physiologie Animale 1-1Document150 pagesCours Physiologie Animale 1-1Mina OussarPas encore d'évaluation
- 8-Tissu NerveuxDocument99 pages8-Tissu NerveuxAy ManPas encore d'évaluation
- 5 - Physiologie Du NeuroneDocument8 pages5 - Physiologie Du Neuronemintchimechak2Pas encore d'évaluation
- Tissus NerveuxDocument19 pagesTissus NerveuxGuenzet Akila100% (1)
- Chap II Tissu NerveuxDocument43 pagesChap II Tissu NerveuxbadamassimarwanePas encore d'évaluation
- 01-Physilogie Du Tissu NerveuxDocument49 pages01-Physilogie Du Tissu NerveuxBook HunterPas encore d'évaluation
- BIOLE102 Neurobiologie: LégendeDocument117 pagesBIOLE102 Neurobiologie: LégendeDaniela RiosPas encore d'évaluation
- Le Système NerveuxDocument8 pagesLe Système NerveuxazizPas encore d'évaluation
- Le Tissu NerveuxDocument2 pagesLe Tissu NerveuxslimdamakPas encore d'évaluation
- Tissu NerveuxDocument36 pagesTissu NerveuxBMA-medecinePas encore d'évaluation
- Partie 4Document31 pagesPartie 4ndeye yarame diopPas encore d'évaluation
- Anatomie Physiologie Du NerfDocument7 pagesAnatomie Physiologie Du NerfMohammed GartitPas encore d'évaluation
- La Cellule Nerveuse W.PDF KKDocument6 pagesLa Cellule Nerveuse W.PDF KKas.cherfaouiPas encore d'évaluation
- Ganglion Nerveux FinalDocument3 pagesGanglion Nerveux Finalmalakzerizer2005Pas encore d'évaluation
- Tissu Nerveux Partie 1Document93 pagesTissu Nerveux Partie 1Yaya TraoréPas encore d'évaluation
- 3-Le Tissu NerveuxDocument19 pages3-Le Tissu Nerveuxboutefal imenePas encore d'évaluation
- Sintesis 126 + Capitulo 11Document133 pagesSintesis 126 + Capitulo 11Daniela RiosPas encore d'évaluation
- CM4 - Vascularisation Et Voies Sensitivo-MotricesDocument14 pagesCM4 - Vascularisation Et Voies Sensitivo-Motricescharbelantoun2443Pas encore d'évaluation
- Neurophysiologie(1)Document15 pagesNeurophysiologie(1)aitbaali.oriflamePas encore d'évaluation
- L'oeilDocument8 pagesL'oeilmalakzerizer2005Pas encore d'évaluation
- Physiologie Animale Cours 7Document35 pagesPhysiologie Animale Cours 7Alhassane BahPas encore d'évaluation
- NeurobiologieDocument11 pagesNeurobiologietinaPas encore d'évaluation
- Histologie SNCDocument7 pagesHistologie SNCSuzanne BorelPas encore d'évaluation
- Neuroanat-2 0Document83 pagesNeuroanat-2 0seloua kaid100% (1)
- Embryologie NerveuxDocument6 pagesEmbryologie Nerveuxmalakzerizer2005Pas encore d'évaluation
- 8 Tissu NerveuxDocument11 pages8 Tissu NerveuxEddy ChahedPas encore d'évaluation
- Chapitre 3 Le Système NerveuxDocument26 pagesChapitre 3 Le Système NerveuxdfhohfeicxhPas encore d'évaluation
- SYSTEME NERVEUX RechDocument14 pagesSYSTEME NERVEUX Rechariane.vasseur29Pas encore d'évaluation
- Le Système Nerveux PDFDocument13 pagesLe Système Nerveux PDFSaggou OuafaPas encore d'évaluation
- UE2 - Histologie - Tissu NerveuxDocument38 pagesUE2 - Histologie - Tissu Nerveuxsun-nee-chan9Pas encore d'évaluation
- Neurobiologie: Les Grands Articles d'UniversalisD'EverandNeurobiologie: Les Grands Articles d'UniversalisPas encore d'évaluation
- Les VitaminesDocument13 pagesLes VitaminesBemmoussat AsmaPas encore d'évaluation
- MECANISMES DABSORPTION ET MOUVEMENT DES IONS KaidDocument5 pagesMECANISMES DABSORPTION ET MOUVEMENT DES IONS KaidBemmoussat AsmaPas encore d'évaluation
- Betterave FourragereDocument85 pagesBetterave FourragereBemmoussat AsmaPas encore d'évaluation
- L'étude Agro-Pédologique D'un Sol: Une Sortie Le But de La SortieDocument3 pagesL'étude Agro-Pédologique D'un Sol: Une Sortie Le But de La SortieBemmoussat AsmaPas encore d'évaluation
- Vigne de Table PowerDocument24 pagesVigne de Table PowerBemmoussat AsmaPas encore d'évaluation
- Aubergine WordDocument5 pagesAubergine WordBemmoussat Asma100% (1)
- 2 - Le Tissu CartilagineuxDocument5 pages2 - Le Tissu CartilagineuxBemmoussat AsmaPas encore d'évaluation
- 06-Les Tissus MusculairesDocument9 pages06-Les Tissus MusculairesBemmoussat AsmaPas encore d'évaluation
- TD N°1 en Biologie Moléculaire Et Génie GénétiqueDocument4 pagesTD N°1 en Biologie Moléculaire Et Génie GénétiqueBemmoussat AsmaPas encore d'évaluation
- Glossaire GénétiqueDocument13 pagesGlossaire GénétiqueBemmoussat AsmaPas encore d'évaluation
- Cours1 Neuro1Document19 pagesCours1 Neuro1Eya MoknessiPas encore d'évaluation
- NeurohistologieDocument52 pagesNeurohistologiemeriemPas encore d'évaluation
- Tissus NerveuxDocument19 pagesTissus NerveuxGuenzet Akila100% (1)
- Exercices de SVT Le Fonctionnement Des Neurones 1Document12 pagesExercices de SVT Le Fonctionnement Des Neurones 1soilihi ibnamriPas encore d'évaluation
- Le Tissu NerveuxDocument51 pagesLe Tissu NerveuxYouness KhalfaouiPas encore d'évaluation
- Tissu Nerveux Partie 1Document93 pagesTissu Nerveux Partie 1Yaya TraoréPas encore d'évaluation
- Tableau NeurophysiologieDocument2 pagesTableau NeurophysiologieSAMI DHAOUIPas encore d'évaluation
- EvaluationsDocument6 pagesEvaluationsbayaPas encore d'évaluation
- Tissu NerveuxDocument36 pagesTissu Nerveuxfederica.lamaPas encore d'évaluation
- CM 1 Neurophysiologie - Introduction Et Cellules Gliales N GuéroutDocument82 pagesCM 1 Neurophysiologie - Introduction Et Cellules Gliales N GuéroutfezbejfPas encore d'évaluation
- GVVGKDocument5 pagesGVVGKNiangPas encore d'évaluation
- Cours SVT Tle D Ecole OnlineDocument188 pagesCours SVT Tle D Ecole OnlineEnfant de Dieu KidsPas encore d'évaluation
- MOD6 Le Syste Me Nerveux Central - La Moelle e Pinie Re PDFDocument3 pagesMOD6 Le Syste Me Nerveux Central - La Moelle e Pinie Re PDFmedPas encore d'évaluation
- Le Tissu Nerveux - Vue GénéraleDocument67 pagesLe Tissu Nerveux - Vue GénéraleSiaka YodaPas encore d'évaluation
- 6404c3c29c6ef - Révision Générale Pour Le DS2 4MDocument5 pages6404c3c29c6ef - Révision Générale Pour Le DS2 4MBoruto UzumakiPas encore d'évaluation
- TD 1 PhyADocument7 pagesTD 1 PhyAMamadou lamine DiattaPas encore d'évaluation
- NerveuxDocument32 pagesNerveuxMinou Meha100% (2)
- Physio 1Document3 pagesPhysio 1AbdelOuahidSenhadjiPas encore d'évaluation
- Tissu Nerveux - PolycopiéDocument10 pagesTissu Nerveux - PolycopiéAmina DrkPas encore d'évaluation
- Systeme NerveuxDocument42 pagesSysteme NerveuxMERIEM RIMPas encore d'évaluation
- Le Cortex Cérébelleux Partie 2Document9 pagesLe Cortex Cérébelleux Partie 2louanne kloepferPas encore d'évaluation
- Cours de Biologie en Ligne S4Document9 pagesCours de Biologie en Ligne S4Jean selondieu100% (1)
- Physiologie Humaine Chap 2: Bases Cellulaires Du Contrôle NerveuxDocument77 pagesPhysiologie Humaine Chap 2: Bases Cellulaires Du Contrôle NerveuxLaurinePas encore d'évaluation
- Le Tissu NerveuxDocument16 pagesLe Tissu NerveuxZizou HamdiPas encore d'évaluation
- Uef 102 Diapo Tissu NerveuxDocument123 pagesUef 102 Diapo Tissu NerveuxAlia YmfPas encore d'évaluation
- Le Tissu NerveuxDocument13 pagesLe Tissu NerveuxManar Bahi100% (1)
- Cours Neurohisto PCEM1 2006-2007Document50 pagesCours Neurohisto PCEM1 2006-2007Wiwi LauraPas encore d'évaluation
- Chapitre III-partie 1Document15 pagesChapitre III-partie 1Yosr LenglizPas encore d'évaluation
- Physiologie Cours COMPLETDocument492 pagesPhysiologie Cours COMPLETProjojo99Pas encore d'évaluation
- NeuroneDocument25 pagesNeuroneSoumaila GanaPas encore d'évaluation