Vous aimerez peut-être aussi
- Management de La Relation Client: Support de Cours Semestre IIDocument18 pagesManagement de La Relation Client: Support de Cours Semestre IIsekkat salmaPas encore d'évaluation
- Fiche Technique 1 - Pannar 12 Et 53 CouleurDocument4 pagesFiche Technique 1 - Pannar 12 Et 53 CouleurHoulpou Domba100% (6)
- Thermal SourceDocument1 pageThermal SourceMehdi JeddiPas encore d'évaluation
- Monographie Wilaya Souk AhrasDocument22 pagesMonographie Wilaya Souk AhrasMack ElevenPas encore d'évaluation
- Données (Etude 2013)Document23 pagesDonnées (Etude 2013)sabahPas encore d'évaluation
- 1 - 2019 - 26 Images HP Et Atlas SaharienDocument18 pages1 - 2019 - 26 Images HP Et Atlas Saharienapi-71796264Pas encore d'évaluation
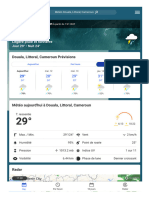
- Douala, Littoral, CamerounConditionsetprévisionsmétéo TheWeatherChannelWeather - Com 1694415002562Document2 pagesDouala, Littoral, CamerounConditionsetprévisionsmétéo TheWeatherChannelWeather - Com 1694415002562Andrea AngelaPas encore d'évaluation
- Classement Générale 11kmDocument3 pagesClassement Générale 11kmCaroline GirardPas encore d'évaluation
- Groupes de Stage 1Document2 pagesGroupes de Stage 1meriemaya AskarPas encore d'évaluation
- TABLEAU CompletDocument1 pageTABLEAU Completmaxou DPas encore d'évaluation
- IntroductionDocument5 pagesIntroductionNangdong Zebaze Ulrich DulainPas encore d'évaluation
- Classement Du 10 KM de l'AJA MarathonDocument8 pagesClassement Du 10 KM de l'AJA MarathonHugo BORRELPas encore d'évaluation
- Orca Share Media1655754734626 6944738706464949087Document5 pagesOrca Share Media1655754734626 6944738706464949087Àbdenöùr SâhràòùiPas encore d'évaluation
- TypeDocument5 pagesTypeWilfried TsanangPas encore d'évaluation
- Séismes Aujourd'hui Derniers Tremblements de TerDocument1 pageSéismes Aujourd'hui Derniers Tremblements de Terirtadmira37Pas encore d'évaluation
- Unicef 230213 01 02Document36 pagesUnicef 230213 01 02Rodolphe MukokoPas encore d'évaluation
- Contamination Bacteriologique Des Eaux Souterraines Par Les Eaux Usees de La Ville de Settat, MarocDocument21 pagesContamination Bacteriologique Des Eaux Souterraines Par Les Eaux Usees de La Ville de Settat, MarocRach Ben100% (1)
- Panneaux 100% Pris en Charge: Mosquée EssalamDocument1 pagePanneaux 100% Pris en Charge: Mosquée EssalamShiraz AmirPas encore d'évaluation
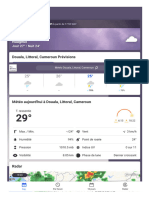
- Douala, Littoral, CamerounConditionsetprévisionsmétéo TheWeatherChannelWeather - Com 1694538343091Document3 pagesDouala, Littoral, CamerounConditionsetprévisionsmétéo TheWeatherChannelWeather - Com 1694538343091Andrea AngelaPas encore d'évaluation
- Outafat AbderrahmaneDocument77 pagesOutafat AbderrahmanearachidPas encore d'évaluation
- Classement Générale 22kmDocument2 pagesClassement Générale 22kmCaroline GirardPas encore d'évaluation
- Senegal NdeneDocument15 pagesSenegal NdeneSATFSCRIBPas encore d'évaluation
- Echium Modestum, Plantago Benisnassenii, Teucrium Doumerguei, Nouveaux Taxons Pour La Flore D'algérieDocument16 pagesEchium Modestum, Plantago Benisnassenii, Teucrium Doumerguei, Nouveaux Taxons Pour La Flore D'algériesekkalPas encore d'évaluation
- Liste Onmc 2022-12-14Document66 pagesListe Onmc 2022-12-14gromologieee gromoPas encore d'évaluation
- Classement Générale Marche NordiqueDocument1 pageClassement Générale Marche NordiqueCaroline GirardPas encore d'évaluation
- Midpoint Online Calculator, Astrology Midpoints, Cosmobiology - PDFDocument4 pagesMidpoint Online Calculator, Astrology Midpoints, Cosmobiology - PDFThe ManPas encore d'évaluation
- RdsDocument4 pagesRdsWilfried TsanangPas encore d'évaluation
- CHAPITRE 02 LES AVTIVITE SOCIALES Wilaya de BatnaDocument66 pagesCHAPITRE 02 LES AVTIVITE SOCIALES Wilaya de BatnaAïchaPas encore d'évaluation
- Présentation Du TangerDocument2 pagesPrésentation Du Tangerelmeskini marowaPas encore d'évaluation
- Preturi C5 C6 Fara Tva August 2023Document2 pagesPreturi C5 C6 Fara Tva August 2023paraipanusPas encore d'évaluation
- Etourneau - Viaduc de Barbin Circuit À Partir de Saint-Laurent-sur-Sèvre Randonnée Pédestre KomootDocument3 pagesEtourneau - Viaduc de Barbin Circuit À Partir de Saint-Laurent-sur-Sèvre Randonnée Pédestre KomootYannickPas encore d'évaluation
- Solar AmitDocument10 pagesSolar AmitAbdelilah HilaliPas encore d'évaluation
- Fiche Signaletique P7Document1 pageFiche Signaletique P7sadatt5Pas encore d'évaluation
- Fiche Signaletique P8Document1 pageFiche Signaletique P8sadatt5Pas encore d'évaluation
- Manuel D'utilisation: Systeme D'Information Geographique Sur Les Oasis en MauritanieDocument54 pagesManuel D'utilisation: Systeme D'Information Geographique Sur Les Oasis en MauritanieGeorges Patrick MANDENGPas encore d'évaluation
- 554a4ac3-f591-41ef-8a09-d3b0829f7a1cDocument101 pages554a4ac3-f591-41ef-8a09-d3b0829f7a1cprachedes carlosPas encore d'évaluation
- Rapport Géophysique Tanandava Station Madagascar 2015Document25 pagesRapport Géophysique Tanandava Station Madagascar 2015ChristinahPas encore d'évaluation
- SpeachDocument17 pagesSpeachra-harinanjaPas encore d'évaluation
- Météo Agadir 14 Jours - Tameteo - Com MeteoredDocument1 pageMétéo Agadir 14 Jours - Tameteo - Com MeteoredSelrahc NPas encore d'évaluation
- Rapport Autorisation D Exploration Diaban Sud 2020Document33 pagesRapport Autorisation D Exploration Diaban Sud 2020Oumar KonePas encore d'évaluation
- ST S1 2019 2020 SemestreDocument112 pagesST S1 2019 2020 SemestreAmmar GouasmiaPas encore d'évaluation
- 2.2 Les Régions: Voir La CarteDocument9 pages2.2 Les Régions: Voir La CarteMohamedSociologiePas encore d'évaluation
- POA TB Paru PKM KPDocument13 pagesPOA TB Paru PKM KPAde Surya DarmaPas encore d'évaluation
- فــي طــوبــىDocument1 pageفــي طــوبــىMaguette SeckPas encore d'évaluation
- 05 DatumProjections 2012Document59 pages05 DatumProjections 2012Mahad AbdiPas encore d'évaluation
- Home Maintenant Temps Horaire Aujourd'hui: AdvertisementsDocument1 pageHome Maintenant Temps Horaire Aujourd'hui: AdvertisementsbenbelkacemPas encore d'évaluation
- 6 - 2021 - 126 Images HP Et Atlas SaharienDocument60 pages6 - 2021 - 126 Images HP Et Atlas Saharienapi-71796264Pas encore d'évaluation
- Rapport N°15729316Document3 pagesRapport N°15729316إعلام الآليPas encore d'évaluation
- GENDARMERIEDocument2 pagesGENDARMERIEGATTA michelPas encore d'évaluation
- AnalyseDocument44 pagesAnalyseMiracle L'amourePas encore d'évaluation
- Piece Ecrite s1 PlanificationDocument9 pagesPiece Ecrite s1 PlanificationIbtissem BensalahPas encore d'évaluation
- BLCKEDDocument7 pagesBLCKEDCélicole Kaïs MakasiPas encore d'évaluation
- Analyse Demande de transportIII.2Document8 pagesAnalyse Demande de transportIII.2yestephane2Pas encore d'évaluation
- PRO 12 Procedure Accreditation Rev12!17!01 2023Document13 pagesPRO 12 Procedure Accreditation Rev12!17!01 2023nibelnina0Pas encore d'évaluation
- Cours SurvieDocument59 pagesCours SurvieFranck BouePas encore d'évaluation
- Pièces Détachées Cuisinière SiemensDocument2 pagesPièces Détachées Cuisinière SiemensSaattajaPas encore d'évaluation
- Colloque2019 b105 Confortement Des Barrages en Maconnerie Par Des Recharges Aval en Remblai Ou en EnrochementsDocument12 pagesColloque2019 b105 Confortement Des Barrages en Maconnerie Par Des Recharges Aval en Remblai Ou en EnrochementsMohamed Zakaria ZaidiPas encore d'évaluation
- Vers Une Stratégie de Gestion Intégrée Des Risques Au MarocDocument24 pagesVers Une Stratégie de Gestion Intégrée Des Risques Au MarocINSAF ELYAHYAOUIPas encore d'évaluation
- PFE DROIT DE LA CONCURRENCE Et PME-1Document175 pagesPFE DROIT DE LA CONCURRENCE Et PME-1meryemlaf1996Pas encore d'évaluation
- ECS2-05-Série Dexercices N°5 - Statistique Exhaustive Et Information de Fisher - CorrigéDocument7 pagesECS2-05-Série Dexercices N°5 - Statistique Exhaustive Et Information de Fisher - CorrigéikhlasPas encore d'évaluation
- Etude de Cas Loi Exponentielle 2021Document12 pagesEtude de Cas Loi Exponentielle 2021Fatima DaoudiPas encore d'évaluation
- ENTREPRENARITDocument121 pagesENTREPRENARITCédric MaatheyPas encore d'évaluation
- Cours Management Staratégique 202 2Document54 pagesCours Management Staratégique 202 2Abdessamad Elguezzam100% (1)
- Travailler Avec EviewsDocument25 pagesTravailler Avec EviewsHatim El OtmaniPas encore d'évaluation
- La Rse Dans Les PmeDocument11 pagesLa Rse Dans Les PmeHamza MechehatPas encore d'évaluation
- Examen Final de Stat Compt en LigneDocument4 pagesExamen Final de Stat Compt en LigneLOKOSSOU DURANDPas encore d'évaluation
- Pfe FinaaaaalDocument71 pagesPfe Finaaaaalmouhcine berarePas encore d'évaluation
- Roland Berger Transformation DigitaleDocument27 pagesRoland Berger Transformation DigitaleL'Usine NouvellePas encore d'évaluation
- Gomes Romeu DiffDocument301 pagesGomes Romeu Diffia2e techPas encore d'évaluation
- NV PDTDocument16 pagesNV PDTYousra EssammaâPas encore d'évaluation
- PNDS Francais Final 12.07.2017Document148 pagesPNDS Francais Final 12.07.2017Germain LumaPas encore d'évaluation
- 1 PBDocument16 pages1 PBEl Ghandour Fatima EzzahraePas encore d'évaluation
- TP Statistiques DescriptivesDocument2 pagesTP Statistiques DescriptivesYou BgPas encore d'évaluation
- Doctorant À l'ENSSEA, Maitre Assistant Classe À l'ENPDocument29 pagesDoctorant À l'ENSSEA, Maitre Assistant Classe À l'ENPsiwarPas encore d'évaluation
- Rapport D'échange - FR-Pierre-Eloi-Gay-FGV-EAESPDocument5 pagesRapport D'échange - FR-Pierre-Eloi-Gay-FGV-EAESPPierre Eloi GayPas encore d'évaluation
- Exercice AgrégatDocument2 pagesExercice AgrégatELAMRANIYOUSSEF100% (6)
- Rapport - Vers Une Économie de La Fonctionnalité À Haute Valeur Environnementale Et Sociale en 2050Document300 pagesRapport - Vers Une Économie de La Fonctionnalité À Haute Valeur Environnementale Et Sociale en 2050Patrice LEBONPas encore d'évaluation