Vous aimerez peut-être aussi
- Arobas5 - t8 - Texte - 8 - L'homme Aux Quarante Manteaux CorrDocument4 pagesArobas5 - t8 - Texte - 8 - L'homme Aux Quarante Manteaux CorrCassandra Maalouf100% (1)
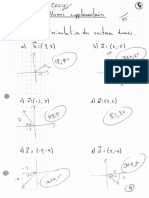
- Ex.6 - CorrigéDocument14 pagesEx.6 - CorrigéCassandra MaaloufPas encore d'évaluation
- Entente Sur La Sécurité Au LaboratoireDocument1 pageEntente Sur La Sécurité Au LaboratoireCassandra MaaloufPas encore d'évaluation
- Ex. 7 - CorrigéDocument14 pagesEx. 7 - CorrigéCassandra MaaloufPas encore d'évaluation
- #3 Le Parc Marin Du Saguenay CorrigéDocument4 pages#3 Le Parc Marin Du Saguenay CorrigéCassandra MaaloufPas encore d'évaluation
- H2022 - Misession 1 - CorrigéDocument8 pagesH2022 - Misession 1 - CorrigéCassandra MaaloufPas encore d'évaluation
- Ex. 5 - CorrigéDocument7 pagesEx. 5 - CorrigéCassandra MaaloufPas encore d'évaluation
- Expérience 1: Chromatographie Sur Couche MinceDocument14 pagesExpérience 1: Chromatographie Sur Couche MinceCassandra MaaloufPas encore d'évaluation
- Smart Biolog I eDocument7 pagesSmart Biolog I eCassandra MaaloufPas encore d'évaluation
- Exercices Supplémentaires PHY 5042Document124 pagesExercices Supplémentaires PHY 5042Cassandra MaaloufPas encore d'évaluation
- Expérience 2: Purification de Produits Chimiques Par DistillationDocument15 pagesExpérience 2: Purification de Produits Chimiques Par DistillationCassandra MaaloufPas encore d'évaluation
- CHI-5061 Prétestb CsdgsDocument9 pagesCHI-5061 Prétestb CsdgsCassandra Maalouf0% (1)
- Devoir 1 - Unité 1 (BS) 2023Document8 pagesDevoir 1 - Unité 1 (BS) 2023Cassandra MaaloufPas encore d'évaluation
- La CelluleDocument4 pagesLa CelluleCassandra MaaloufPas encore d'évaluation
- CD1Lesauvetage (Exp, Valeurabsolue, Contraintes)Document6 pagesCD1Lesauvetage (Exp, Valeurabsolue, Contraintes)Cassandra MaaloufPas encore d'évaluation
- Cours 5-6 NotesDocument26 pagesCours 5-6 NotesCassandra MaaloufPas encore d'évaluation
- Os Chimie Chap1 p59-62 Exercices SyntheseDocument4 pagesOs Chimie Chap1 p59-62 Exercices SyntheseCassandra MaaloufPas encore d'évaluation
- Os Chimie Chap1 p46-49 ExercicesDocument4 pagesOs Chimie Chap1 p46-49 ExercicesCassandra MaaloufPas encore d'évaluation
- Revision Angles CDocument4 pagesRevision Angles CCassandra MaaloufPas encore d'évaluation
- Bio Review Exam1Document2 pagesBio Review Exam1Cassandra MaaloufPas encore d'évaluation
- Chimie Chap0 p21-32Document14 pagesChimie Chap0 p21-32Cassandra Maalouf100% (1)
- Revision Angles CDocument4 pagesRevision Angles CCassandra MaaloufPas encore d'évaluation
- Os Chimie Chap1 p39-40 ExercicesDocument2 pagesOs Chimie Chap1 p39-40 ExercicesCassandra MaaloufPas encore d'évaluation
- Revision Angles CDocument4 pagesRevision Angles CCassandra MaaloufPas encore d'évaluation
- Revisionfinale Math068 536 8Document8 pagesRevisionfinale Math068 536 8Cassandra MaaloufPas encore d'évaluation
- Résumer 1Document4 pagesRésumer 1Cassandra MaaloufPas encore d'évaluation
- Orientation VecteurDocument2 pagesOrientation VecteurCassandra MaaloufPas encore d'évaluation
- Les ConiquesDocument2 pagesLes ConiquesCassandra MaaloufPas encore d'évaluation
- Notes Sur Les Enfants Confrontées A Deux CulturesDocument1 pageNotes Sur Les Enfants Confrontées A Deux CulturesCassandra MaaloufPas encore d'évaluation
- Algorithme Et Les Instructions de Base + ConditionelleDocument12 pagesAlgorithme Et Les Instructions de Base + ConditionelleAGUERJA DOUHA100% (1)
- 2015 td2 PDFDocument4 pages2015 td2 PDFأيمن أيمنPas encore d'évaluation
- Betonb 30Document12 pagesBetonb 30Mandjou Boune Saïd FofanaPas encore d'évaluation
- Pfe Meryem SlimaniDocument78 pagesPfe Meryem SlimaniMed MohamedPas encore d'évaluation
- Structures Statistiques Et EstimationDocument107 pagesStructures Statistiques Et EstimationanasrachidPas encore d'évaluation
- MiCOM P341 - 1124271196185-P341-922g - FR - 1150Document8 pagesMiCOM P341 - 1124271196185-P341-922g - FR - 1150Polux LefebvrePas encore d'évaluation
- Cours 7 - SuitDocument4 pagesCours 7 - SuitMadjidAvengersPas encore d'évaluation
- Bases Linux S22Document12 pagesBases Linux S22Yan MEKUEKOPas encore d'évaluation
- Fiches Methodes Cycle 4Document39 pagesFiches Methodes Cycle 4PELLISSIERPas encore d'évaluation
- Exos Polytechnique Mécanique QuantiqueDocument78 pagesExos Polytechnique Mécanique QuantiqueMath055100% (1)
- La Programmation Linéaire EXERCICES CORRIGESDocument32 pagesLa Programmation Linéaire EXERCICES CORRIGESahlem100% (2)
- ABB Technical Guide No 1 REVCDocument72 pagesABB Technical Guide No 1 REVCdrevetph6233Pas encore d'évaluation
- Corrections Des Exercices Du TP N°1: Infos Système Uname - ADocument14 pagesCorrections Des Exercices Du TP N°1: Infos Système Uname - AradouanePas encore d'évaluation
- Chapitre 12Document4 pagesChapitre 12trikiPas encore d'évaluation
- 2-Profil en LongDocument30 pages2-Profil en LongfrancoPas encore d'évaluation
- MLCCPLMXDocument17 pagesMLCCPLMXBra. DekPas encore d'évaluation
- Exercices Corriges Suites ReellesDocument28 pagesExercices Corriges Suites ReellesMCHICHI ALAMIPas encore d'évaluation
- Axe-FX-II Manual FRDocument211 pagesAxe-FX-II Manual FRFouquetPas encore d'évaluation
- RATIONALISMEDocument5 pagesRATIONALISMEreineesther941Pas encore d'évaluation
- Vest 52137653Document29 pagesVest 52137653AINA ANDRIAMAHENIKAJATSIALONINAPas encore d'évaluation
- Access 2007 Et VBADocument153 pagesAccess 2007 Et VBADominiquePas encore d'évaluation
- Formalisation Et Résolution Des Problèmes de Découpes LinéairesDocument19 pagesFormalisation Et Résolution Des Problèmes de Découpes LinéairesxavierPas encore d'évaluation
- Corail Catalogue Général 2021 PDFDocument325 pagesCorail Catalogue Général 2021 PDFBla BlePas encore d'évaluation
- Amdec: AMDEC: Analyse Des Modes de Défaillances, deDocument13 pagesAmdec: AMDEC: Analyse Des Modes de Défaillances, deRim BdidaPas encore d'évaluation
- Masse de TerreDocument5 pagesMasse de Terreأبو رؤوف رابح شرياقPas encore d'évaluation
- TD Microcontroleur 2Document25 pagesTD Microcontroleur 2Ismael GhaziPas encore d'évaluation
- Capture D'écran . 2021-07-01 À 16.00.17Document10 pagesCapture D'écran . 2021-07-01 À 16.00.17Souhail BenbrahimPas encore d'évaluation
- TechnologieDocument8 pagesTechnologieSamir RimasPas encore d'évaluation
- Cours RC2 Séance N°2 (RIP)Document38 pagesCours RC2 Séance N°2 (RIP)Fatima MouroPas encore d'évaluation
- Utilisation Catia Brep FRDocument4 pagesUtilisation Catia Brep FRBen FahPas encore d'évaluation