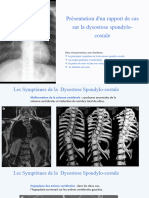
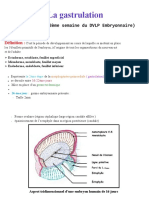
Vous aimerez peut-être aussi
- Resume Mollusques 2Document2 pagesResume Mollusques 2DINO ECOPas encore d'évaluation
- Resume Mollusques 1Document2 pagesResume Mollusques 1Ny Aina AndriamaroPas encore d'évaluation
- Ensb Lessons-Embryo OursinDocument28 pagesEnsb Lessons-Embryo Oursinsirine falehPas encore d'évaluation
- Les MollusquesDocument222 pagesLes Mollusquesshanksgamer22Pas encore d'évaluation
- Chapitre 4 - Les MollusquesDocument14 pagesChapitre 4 - Les MollusquesLopezz GodwinoPas encore d'évaluation
- Les EchinodermesDocument23 pagesLes EchinodermeslouloulesboisPas encore d'évaluation
- TD L2 Biologie de La Reproduction 2019Document29 pagesTD L2 Biologie de La Reproduction 2019ottey etienne yoroPas encore d'évaluation
- Cours Biologie Marine Vers PDFDocument7 pagesCours Biologie Marine Vers PDFAnonymous PKVCsGPas encore d'évaluation
- Prsentationfcondation3 140503113639 Phpapp01 PDFDocument99 pagesPrsentationfcondation3 140503113639 Phpapp01 PDFModeste RAKOTOARIMANANAPas encore d'évaluation
- Cours Biologie Marine Bryozoaires Ligne PDFDocument6 pagesCours Biologie Marine Bryozoaires Ligne PDFAnonymous PKVCsGPas encore d'évaluation
- STM Ani ReimersDocument172 pagesSTM Ani ReimersTony PobranPas encore d'évaluation
- Exposé MammifèresDocument4 pagesExposé Mammifèressteve fotsoPas encore d'évaluation
- La Reproduction Sexuee Chez Les Plantes Sans Fleurs Cours 1Document10 pagesLa Reproduction Sexuee Chez Les Plantes Sans Fleurs Cours 1youness kmiraPas encore d'évaluation
- Zoologie 2 DidermiquesDocument56 pagesZoologie 2 Didermiquessabdo3991Pas encore d'évaluation
- Préparé Par M. SOUTTOU K. Chapitre 6: Embranchement VertébrésDocument4 pagesPréparé Par M. SOUTTOU K. Chapitre 6: Embranchement VertébrésIbrahim Mahamat AminamiPas encore d'évaluation
- Explorer Annexe EscargotsDocument4 pagesExplorer Annexe EscargotsFaycalHamamaPas encore d'évaluation
- Ctenaires PDFDocument13 pagesCtenaires PDFNsangwaPas encore d'évaluation
- Bryozoaires 2023Document10 pagesBryozoaires 2023kondo fatoumataPas encore d'évaluation
- EchinodermesDocument8 pagesEchinodermesCyrine. ChPas encore d'évaluation
- Cours Biologie Marine Mollusques PPT Ligne PDFDocument8 pagesCours Biologie Marine Mollusques PPT Ligne PDFAnonymous PKVCsGPas encore d'évaluation
- Mollusques StudDocument29 pagesMollusques StudSwayli HossamPas encore d'évaluation
- ChapI Annexes Emb Controle DevDocument6 pagesChapI Annexes Emb Controle DevBabel BodjonaPas encore d'évaluation
- WP Contentuploads201111cours Biologie Marine Mollusques PPT Ligne PDFDocument8 pagesWP Contentuploads201111cours Biologie Marine Mollusques PPT Ligne PDFCeyda BAŞ100% (1)
- Corrigé TD1 Zoologie Invertebres Juin2022Document8 pagesCorrigé TD1 Zoologie Invertebres Juin2022elhadjousmanesambe46Pas encore d'évaluation
- Chapitre 4 Bio AnDocument10 pagesChapitre 4 Bio AndelphinemorangePas encore d'évaluation
- MollusquesDocument81 pagesMollusquesYasmine BoujerfaouiPas encore d'évaluation
- Chapitre 6 Les Ostracodes VivantsDocument9 pagesChapitre 6 Les Ostracodes VivantsjahyokotaPas encore d'évaluation
- Examen4 Biologie Animales SV2Document2 pagesExamen4 Biologie Animales SV2Doudou AminePas encore d'évaluation
- Deutérostomiens1 2023-2024Document47 pagesDeutérostomiens1 2023-2024marammari40Pas encore d'évaluation
- Cours Bio Animale 2 Des Parazoaires Aux Protostomiens (Chapi 4 Et 5)Document17 pagesCours Bio Animale 2 Des Parazoaires Aux Protostomiens (Chapi 4 Et 5)alexmount321Pas encore d'évaluation
- Biodiversité Animale 2 - Les EchinodermesDocument3 pagesBiodiversité Animale 2 - Les EchinodermesMarine CondezPas encore d'évaluation
- BIOLOGIE1TD3groupes3 4 5et6L1Geo21Document4 pagesBIOLOGIE1TD3groupes3 4 5et6L1Geo21aqua FishPas encore d'évaluation
- Anatomie D'un Escargot (Schéma en Coupe) : Schéma Extrait de Wikipedia (Libre de Droit)Document2 pagesAnatomie D'un Escargot (Schéma en Coupe) : Schéma Extrait de Wikipedia (Libre de Droit)kpegoh komlanPas encore d'évaluation
- Cours Cnidaires Et CténairesDocument15 pagesCours Cnidaires Et Cténairesamina imenePas encore d'évaluation
- Evaluation - Examens Finaux - Examen Final 2000 (Corrigé)Document11 pagesEvaluation - Examens Finaux - Examen Final 2000 (Corrigé)mounaPas encore d'évaluation
- Bio Ani Polycopie 1Document13 pagesBio Ani Polycopie 1maomaochongPas encore d'évaluation
- B.A Polycopie 3Document151 pagesB.A Polycopie 3imane chakibPas encore d'évaluation
- SVT2 PolycopiéDocument57 pagesSVT2 Polycopiékhaoula.banouissPas encore d'évaluation
- 9782403041422Document31 pages9782403041422Gedeon Jordan100% (1)
- MétazoairesDocument7 pagesMétazoairesfouad abdelhamidPas encore d'évaluation
- PrésentationDocument18 pagesPrésentationAbou Bakr0% (1)
- Arthropodologie 8Document43 pagesArthropodologie 8Ismail OuskarPas encore d'évaluation
- TP B15 16. Reproduction Et DeveloppementDocument8 pagesTP B15 16. Reproduction Et Developpementjacques yiborPas encore d'évaluation
- Syllabus Unamur ArthropodesDocument55 pagesSyllabus Unamur ArthropodesDINO ECOPas encore d'évaluation
- Chapitre 11 Reproduction Des Oiseaux Embryologie A1 Djeffal PDFDocument14 pagesChapitre 11 Reproduction Des Oiseaux Embryologie A1 Djeffal PDFVéto SamPas encore d'évaluation
- 3g EpongesDocument7 pages3g Epongesramen masterPas encore d'évaluation
- Cours Biologie Marine Mollusques PPT LigneDocument8 pagesCours Biologie Marine Mollusques PPT LigneBabbel KITIOPas encore d'évaluation
- Resume Cours Spongiares Cnidaires Ctenaires 2021Document52 pagesResume Cours Spongiares Cnidaires Ctenaires 2021Anaïs BlPas encore d'évaluation
- TD ParasitologieDocument76 pagesTD ParasitologieSalma Babour100% (1)
- Deutérostomiens2 2023-2024Document44 pagesDeutérostomiens2 2023-2024marammari40Pas encore d'évaluation
- lp1 Ocea 1Document48 pageslp1 Ocea 1belzmar7Pas encore d'évaluation
- BOTANIQUE Chap 4Document27 pagesBOTANIQUE Chap 4Sirandou DiaoPas encore d'évaluation
- Cours 4 Faunistique Spong Cnid Plat Roky 2022Document60 pagesCours 4 Faunistique Spong Cnid Plat Roky 2022Abdelhadi Mehdi ZaatariPas encore d'évaluation
- OVA - C8 - Arthropodes PDF.CDocument23 pagesOVA - C8 - Arthropodes PDF.CNabil holmes100% (1)
- BIO 224 Anatomie - Comparée (2020) - Compressé - 0 PDFDocument81 pagesBIO 224 Anatomie - Comparée (2020) - Compressé - 0 PDFTONDI ABDOU CHACOUR67% (3)
- 4-Cours CnidairesDocument32 pages4-Cours CnidairesAchouri ZouaouiPas encore d'évaluation
- Drag PDFDocument40 pagesDrag PDFDJIL BENPas encore d'évaluation
- Faunistique Cours 11Document14 pagesFaunistique Cours 11ghislain AMEOHAPas encore d'évaluation
- Microsoft Word - Suite Chapitre IIIDocument21 pagesMicrosoft Word - Suite Chapitre IIImanarhajji2024Pas encore d'évaluation
- Appareil génital: Les Grands Articles d'UniversalisD'EverandAppareil génital: Les Grands Articles d'UniversalisPas encore d'évaluation
- Embryologie Generale 3Document9 pagesEmbryologie Generale 3Verone FoullaPas encore d'évaluation
- Embryo23 DiapoDocument127 pagesEmbryo23 DiapoAmina ZirarPas encore d'évaluation
- Résumé Des 4 Semaines - La Chronologie Du Développement EmbryonnaireDocument4 pagesRésumé Des 4 Semaines - La Chronologie Du Développement Embryonnairenacim fethy halesPas encore d'évaluation
- Troisième Semaine Du Développement EmbryonnaireDocument60 pagesTroisième Semaine Du Développement EmbryonnaireMohamedBahmedPas encore d'évaluation
- TD N 8 Gastrulation Chez Amphibiens (Support)Document3 pagesTD N 8 Gastrulation Chez Amphibiens (Support)zana adama coulibalyPas encore d'évaluation
- Regulation Et Determination Des Cellules EmbryonnairesDocument60 pagesRegulation Et Determination Des Cellules EmbryonnairesGhiles Anis OuyahiaPas encore d'évaluation
- Embryologie QCMDocument14 pagesEmbryologie QCMAsgaraly RahimPas encore d'évaluation
- Biologie Animale 3eme Trimestre PDFDocument29 pagesBiologie Animale 3eme Trimestre PDFFafi AliPas encore d'évaluation
- Dysosotose Spondylo-CostaleDocument18 pagesDysosotose Spondylo-Costalejeromeneossi271Pas encore d'évaluation
- Amphi BienDocument15 pagesAmphi BienZie adama DiarrassoubaPas encore d'évaluation
- 3emesemaine PpsDocument38 pages3emesemaine PpsClovis Zue EssandonePas encore d'évaluation
- 647m1 EMBRYOLOGIEDocument20 pages647m1 EMBRYOLOGIEKamilia BilalPas encore d'évaluation
- Troisième Semaine Du Développement Embryonnaire GastrulationDocument40 pagesTroisième Semaine Du Développement Embryonnaire GastrulationNathan DeffoPas encore d'évaluation
- Microsoft Word - Suite Chapitre IIIDocument21 pagesMicrosoft Word - Suite Chapitre IIImanarhajji2024Pas encore d'évaluation
- Acc - EmbryologieDocument14 pagesAcc - Embryologiechahdsouf086Pas encore d'évaluation
- BIO 132 Cours Partie ZooDocument48 pagesBIO 132 Cours Partie ZooBhuvanPas encore d'évaluation
- Développement Embryonnaire Des Oiseaux Partie 2Document21 pagesDéveloppement Embryonnaire Des Oiseaux Partie 2GASMI ABDELNOUR100% (1)
- Revisions Du Cours SVI M2 S1 Avec QCEMDocument52 pagesRevisions Du Cours SVI M2 S1 Avec QCEMazzamPas encore d'évaluation
- GastrulationDocument5 pagesGastrulationMinaheta Guy maurelPas encore d'évaluation
- TRAVAUX DIRIGE DE BOA 356 (DR Wankeu)Document11 pagesTRAVAUX DIRIGE DE BOA 356 (DR Wankeu)Staline DonkengPas encore d'évaluation
- Bases Cellulaires Et Moléculaires Du DéveloppementDocument13 pagesBases Cellulaires Et Moléculaires Du DéveloppementDaVinciPas encore d'évaluation
- DéfinitionDocument17 pagesDéfinitionJouda BivaPas encore d'évaluation
- GastrulationDocument25 pagesGastrulationsousou2000Pas encore d'évaluation
- I. DéfinitionDocument2 pagesI. Définitionhammou HoudjedjePas encore d'évaluation
- Cours Biologie Animale SNV l1 2015 Introduction GeneraleDocument4 pagesCours Biologie Animale SNV l1 2015 Introduction GeneraleSabrin Amani100% (1)
- La Segmentation Chez L'oiseauDocument11 pagesLa Segmentation Chez L'oiseauUMagis Itsfx-Itsa AntadyPas encore d'évaluation
- Embryologie s1Document127 pagesEmbryologie s1MeRyem El MejdoubPas encore d'évaluation
- EmbryogénèseDocument5 pagesEmbryogénèseredPas encore d'évaluation
- A. Copie de 4ème Semaine deDocument3 pagesA. Copie de 4ème Semaine deILHEM RAMDA100% (1)
- Develepomment Embryonnaire UsthbDocument22 pagesDevelepomment Embryonnaire UsthbMahieddine BoucekkinePas encore d'évaluation