Vous aimerez peut-être aussi
- Annexe Série 1 Reproduction MasculineDocument8 pagesAnnexe Série 1 Reproduction Masculinemosrati hedia100% (1)
- 20, Monsieur Vilo: Office Recherche Scientifique Et Technique Ou'Fre-Msr Pahis Entomologie Iviedicale VeterinaireDocument38 pages20, Monsieur Vilo: Office Recherche Scientifique Et Technique Ou'Fre-Msr Pahis Entomologie Iviedicale VeterinaireMarie MamekemPas encore d'évaluation
- Arthropodes HexapodesDocument64 pagesArthropodes HexapodesRASOANOMEJANAHARY Marie OliviaPas encore d'évaluation
- Cours Biologie Marine Vers PDFDocument7 pagesCours Biologie Marine Vers PDFAnonymous PKVCsGPas encore d'évaluation
- Les Pièces Buccales Et L'alimentation Des InsectesDocument8 pagesLes Pièces Buccales Et L'alimentation Des Insectesguyjoe243Pas encore d'évaluation
- 1 - Fiches - de - Groupes TP1 (Pour Le Camembert)Document16 pages1 - Fiches - de - Groupes TP1 (Pour Le Camembert)Maxime lorenzoPas encore d'évaluation
- Clé de Determination Des RhyachophilidaeDocument45 pagesClé de Determination Des RhyachophilidaeMichel-Marie Solito de SolisPas encore d'évaluation
- 3 - Anatomie Generale Des InsectesDocument44 pages3 - Anatomie Generale Des InsectesCedina Özil Yaouza Jr.Pas encore d'évaluation
- 2 Les Premières Branches Des Vertébrés PDFDocument6 pages2 Les Premières Branches Des Vertébrés PDFBelhassen RanimPas encore d'évaluation
- Diaporama Sur La Classification Du VivantDocument25 pagesDiaporama Sur La Classification Du VivantSoumam DieselPas encore d'évaluation
- TP Mammifères DIAPO (2018)Document52 pagesTP Mammifères DIAPO (2018)Bouba SeckPas encore d'évaluation
- Guidebiodiversite Criquets Sauterelles GrillonsDocument16 pagesGuidebiodiversite Criquets Sauterelles GrillonsAlycia SafaPas encore d'évaluation
- CM SVT OiseauMammifDocument57 pagesCM SVT OiseauMammifchaimae.cho96Pas encore d'évaluation
- L'Amateur de papillons: Guide pour la chasse, la préparation et la conservationD'EverandL'Amateur de papillons: Guide pour la chasse, la préparation et la conservationPas encore d'évaluation
- hs30 OrthopteresDocument41 pageshs30 OrthopteresAlycia SafaPas encore d'évaluation
- Cours ZoologieDocument21 pagesCours ZoologieAmcic WeeliPas encore d'évaluation
- Resume Cours Procordes 20221841Document20 pagesResume Cours Procordes 20221841laoudi sarahPas encore d'évaluation
- Le Règne AnimalDocument8 pagesLe Règne AnimalNabil holmesPas encore d'évaluation
- LE REGNE ANIMAL 2 - CopieDocument49 pagesLE REGNE ANIMAL 2 - CopieMohamed ROBAKHIPas encore d'évaluation
- BiologieDocument16 pagesBiologieHillahPas encore d'évaluation
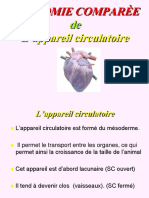
- BIO 224 Anatomie - Comparée (2020) - Compressé - 0 PDFDocument81 pagesBIO 224 Anatomie - Comparée (2020) - Compressé - 0 PDFTONDI ABDOU CHACOUR67% (3)
- Un Regard Sur L Evolution de L HommeDocument6 pagesUn Regard Sur L Evolution de L HommeInés NamPas encore d'évaluation
- BasidioLeucospores PDFDocument6 pagesBasidioLeucospores PDFbrahim chalhoubPas encore d'évaluation
- 01 TaxoDocument13 pages01 Taxoriguelle.lionelPas encore d'évaluation
- AnatomieDocument43 pagesAnatomiezangui hamissou100% (1)
- Biologie Vegetale Cormophytes UjiklDocument44 pagesBiologie Vegetale Cormophytes Ujiklmeryem.bouhamidaPas encore d'évaluation
- VertebresDocument1 pageVertebresBiologie de l'organismePas encore d'évaluation
- Cours 4 Faunistique Spong Cnid Plat Roky 2022Document60 pagesCours 4 Faunistique Spong Cnid Plat Roky 2022Abdelhadi Mehdi ZaatariPas encore d'évaluation
- Entomologie Résumé 02Document9 pagesEntomologie Résumé 02ngoufacktsougmoPas encore d'évaluation
- Pal. Des Vert. 1Document4 pagesPal. Des Vert. 1charaf eddinePas encore d'évaluation
- 3 L'arbre Des Tétrapodes PDFDocument8 pages3 L'arbre Des Tétrapodes PDFBelhassen RanimPas encore d'évaluation
- Licence SV3 Evolution Des EmbryophytesDocument45 pagesLicence SV3 Evolution Des Embryophyteslebonmaelle1Pas encore d'évaluation
- Zoologie Generale FasaDocument92 pagesZoologie Generale FasaRobert Blake100% (1)
- Les MollusquesDocument222 pagesLes Mollusquesshanksgamer22Pas encore d'évaluation
- Faunistique Cours Complet 2021Document69 pagesFaunistique Cours Complet 2021AbdouLlah Ait EdelsPas encore d'évaluation
- 9782403041422Document31 pages9782403041422Gedeon Jordan100% (1)
- Classification - ParasitologieDocument30 pagesClassification - Parasitologiealmnaouar100% (1)
- UntitledDocument10 pagesUntitledJohn LambeauxPas encore d'évaluation
- Araignée TrésorDocument13 pagesAraignée TrésorJean-Marie PrévoteauPas encore d'évaluation
- B.A Les VertebresDocument61 pagesB.A Les Vertebresimane chakibPas encore d'évaluation
- Le Monde VegetalDocument12 pagesLe Monde Vegetalmhamed aadlPas encore d'évaluation
- Cours Anatomie Externe L3 EntomologieDocument188 pagesCours Anatomie Externe L3 Entomologiemohamed rouani100% (1)
- Bilan Arb PhyloDocument4 pagesBilan Arb Phylobeebac2009Pas encore d'évaluation
- Chapitre 1Document21 pagesChapitre 1Sara NailiPas encore d'évaluation
- La Biodiversite Un Instantane Dans L Evolution Du Vivant CorrectionsDocument2 pagesLa Biodiversite Un Instantane Dans L Evolution Du Vivant CorrectionsZiad MattousPas encore d'évaluation
- Cours Botanique 2eme. MycètesDocument50 pagesCours Botanique 2eme. Mycètesbouguessa norelhoudaPas encore d'évaluation
- Algues LPMDocument69 pagesAlgues LPMfiraPas encore d'évaluation
- Partie ApétalesDocument101 pagesPartie ApétalesNassif AnassPas encore d'évaluation
- Cours Botanique 2eme Année. AlguesDocument22 pagesCours Botanique 2eme Année. Alguesbouguessa norelhoudaPas encore d'évaluation
- CoursBA P1Document29 pagesCoursBA P1Yassine Enissay100% (1)
- ArthropodesDocument65 pagesArthropodesmed_yacine100% (1)
- TP 14 Communication Intraspécifique Et Survie de L& 039 EspèceDocument7 pagesTP 14 Communication Intraspécifique Et Survie de L& 039 EspèceErza ScarlettPas encore d'évaluation
- Syllabus 2021 - TP 6Document7 pagesSyllabus 2021 - TP 6adrienbourgeois515Pas encore d'évaluation
- 211120Document16 pages211120Said MessaoudPas encore d'évaluation
- Cours Bio VertébrésDocument24 pagesCours Bio VertébrésClaver PaulPas encore d'évaluation
- Cours Bio Animale 2 Des Parazoaires Aux Protostomiens (Chapi 4 Et 5)Document17 pagesCours Bio Animale 2 Des Parazoaires Aux Protostomiens (Chapi 4 Et 5)alexmount321Pas encore d'évaluation
- Chitinozoaires El Manai Section DDocument23 pagesChitinozoaires El Manai Section DAIT TAHAR NeilaPas encore d'évaluation
- DR Ouattara Ismael Ben Enseignant-Chercheur Ufhb Ufr-Strm Geologueben@Document30 pagesDR Ouattara Ismael Ben Enseignant-Chercheur Ufhb Ufr-Strm Geologueben@KamagatePas encore d'évaluation
- Les Bryophytes - Particularites Et Ecologie - o MannevilleDocument10 pagesLes Bryophytes - Particularites Et Ecologie - o MannevilleRAVELOSON Serge EloiPas encore d'évaluation
- Les InsectesDocument17 pagesLes InsectesFifiMisaPas encore d'évaluation
- HeteropteraDocument19 pagesHeteropteraAli AmroussePas encore d'évaluation
- Fiche 01Document1 pageFiche 01ana414 popoPas encore d'évaluation
- SVT 1Ã Re C - L5 - La Fã©condation Chez Les Mammifã ResDocument11 pagesSVT 1Ã Re C - L5 - La Fã©condation Chez Les Mammifã ResAboulaye DiakitePas encore d'évaluation
- Cours Biof 1Document214 pagesCours Biof 1خديجة الحافةPas encore d'évaluation
- Série 1rep HDocument4 pagesSérie 1rep HWassim Maharsia100% (1)
- L'organisation Generale de L'etre HumainDocument10 pagesL'organisation Generale de L'etre HumainSOLTANIPas encore d'évaluation
- La VermifugationDocument1 pageLa VermifugationBeurier KarinePas encore d'évaluation
- TD3 SujetDocument9 pagesTD3 SujetsafemindPas encore d'évaluation
- Corrige 472944Document117 pagesCorrige 472944DJIL BENPas encore d'évaluation
- BTS Dietetique Le Tissu EpithelialDocument4 pagesBTS Dietetique Le Tissu Epithelialhananeismaili.hiPas encore d'évaluation
- PDF ZootechnieDocument7 pagesPDF ZootechnieGuy Roger OyonoPas encore d'évaluation
- Resume Cours-Zoo-2021-Regne-Animal Et Protozoaire GHDocument104 pagesResume Cours-Zoo-2021-Regne-Animal Et Protozoaire GHAnaïs BlPas encore d'évaluation
- Pauwels Louis - Bergier Jacques - L'Homme Éternel 1Document383 pagesPauwels Louis - Bergier Jacques - L'Homme Éternel 1Materiel2100% (5)
- 5 Cours CtenairesDocument28 pages5 Cours CtenairesAchouri ZouaouiPas encore d'évaluation
- LA DEUXIEME ET LA TROISIEME SEMAINE DE DEVELOPPEMENT COURS Ndiaga L1S2 MédecineDocument89 pagesLA DEUXIEME ET LA TROISIEME SEMAINE DE DEVELOPPEMENT COURS Ndiaga L1S2 MédecineMøüštãphå MëdPas encore d'évaluation
- Immuno 1Document4 pagesImmuno 1Rkia EddabraPas encore d'évaluation
- L'alimentation Du FuturDocument4 pagesL'alimentation Du FuturvaanmonteiroPas encore d'évaluation
- Denice SEMAINE DE LA SCIENCE 2023Document2 pagesDenice SEMAINE DE LA SCIENCE 2023dnicenim99Pas encore d'évaluation
- Asset Upload File521 3873Document940 pagesAsset Upload File521 3873Oumaima AMGHARPas encore d'évaluation
- Comparaison Du Cycle de Développement Chez Le MondeDocument25 pagesComparaison Du Cycle de Développement Chez Le Mondeإلهام فريضيPas encore d'évaluation
- Embranchement Des AnnelidesDocument10 pagesEmbranchement Des AnnelidesNor-eddine CHOUIKHPas encore d'évaluation
- 647m1 EMBRYOLOGIEDocument20 pages647m1 EMBRYOLOGIEKamilia BilalPas encore d'évaluation
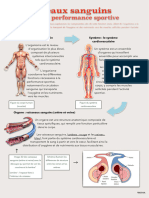
- Appareil CirculatoireDocument39 pagesAppareil CirculatoireCyrine. ChPas encore d'évaluation
- Embryologie Du Systeme Uro-GenitalDocument44 pagesEmbryologie Du Systeme Uro-Genitalsaidbenmokhtar3960Pas encore d'évaluation
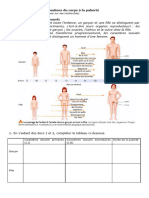
- A1 Puberte RappelsDocument2 pagesA1 Puberte RappelsKim AtuaPas encore d'évaluation
- I. DéfinitionDocument2 pagesI. Définitionhammou HoudjedjePas encore d'évaluation
- 03-Sciences-De-La-Vie-Et-De-La-Terre CM2Document49 pages03-Sciences-De-La-Vie-Et-De-La-Terre CM2Ravaka JuliePas encore d'évaluation
- QCM Histologie Total CorrigeDocument28 pagesQCM Histologie Total Corrigeアブド アマリ92% (13)