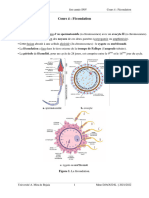
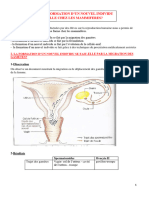
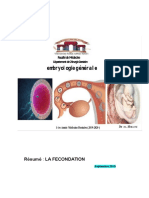
Vous aimerez peut-être aussi
- Chapitre III La FécondationDocument7 pagesChapitre III La FécondationDel HadPas encore d'évaluation
- Cours 4 - Fécondation - Format 2Document7 pagesCours 4 - Fécondation - Format 2Nãřímäñ KŁayPas encore d'évaluation
- Première Semaine Du Développement EmbryonnaireDocument78 pagesPremière Semaine Du Développement EmbryonnaireSofiane BousifePas encore d'évaluation
- H - FécondationDocument25 pagesH - FécondationFotso Nanfa0% (1)
- La Fécondation Cours WordDocument10 pagesLa Fécondation Cours WordNadjib A. NedjarPas encore d'évaluation
- CHAPITRE II Fécondation Mécanismes Cellulaires Et Moléculaires2Document31 pagesCHAPITRE II Fécondation Mécanismes Cellulaires Et Moléculaires2abderrahmanecherchour510Pas encore d'évaluation
- La FecondationDocument23 pagesLa FecondationEmmanuelPas encore d'évaluation
- 0 La Fécondation CoursDocument6 pages0 La Fécondation Coursrolemodel663Pas encore d'évaluation
- 1 Semaine de La FécondationDocument18 pages1 Semaine de La FécondationFofana LancinéPas encore d'évaluation
- Aspect Cellulaire de DéveloppementDocument9 pagesAspect Cellulaire de DéveloppementOm Nawfal NawfalPas encore d'évaluation
- Embryo1an-1ere Semaine2021Document42 pagesEmbryo1an-1ere Semaine2021semep chlefPas encore d'évaluation
- 7 La Premiã Re Semaine Du Dã©veloppement Embryonnaire PPTDocument13 pages7 La Premiã Re Semaine Du Dã©veloppement Embryonnaire PPThamihenbasmaPas encore d'évaluation
- Biologie AnimaleDocument31 pagesBiologie AnimaleBah KhalilPas encore d'évaluation
- A. Copie de 1ère Semaine deDocument3 pagesA. Copie de 1ère Semaine deILHEM RAMDAPas encore d'évaluation
- C 14 F - G - L: HapitreDocument26 pagesC 14 F - G - L: HapitreSouftie ReplayPas encore d'évaluation
- La FecondationDocument13 pagesLa Fecondationmalika mikoPas encore d'évaluation
- F Condation-1e SemDevEmbry PDFDocument22 pagesF Condation-1e SemDevEmbry PDFsoufiane soufPas encore d'évaluation
- Cours 4 - Fécondation - Format 1Document15 pagesCours 4 - Fécondation - Format 1Nãřímäñ KŁayPas encore d'évaluation
- GrosesseDocument12 pagesGrosesseluana.brunelli27Pas encore d'évaluation
- FécondationDocument12 pagesFécondationdyhp2pswsrPas encore d'évaluation
- Cours 4 La FécondationDocument51 pagesCours 4 La FécondationMįSs BęãüTÿPas encore d'évaluation
- Embryo23 05-2mois Devellopement PDFDocument30 pagesEmbryo23 05-2mois Devellopement PDFChouaib MeraoumiaPas encore d'évaluation
- Embryologie Résumé Imane PDFDocument10 pagesEmbryologie Résumé Imane PDFAsmaa ElgattaiaPas encore d'évaluation
- Embryologie Partie1Document69 pagesEmbryologie Partie1Mariame Chakir100% (3)
- HE Fécondation Fichier Produit 1602Document13 pagesHE Fécondation Fichier Produit 1602ZoulkiffiPas encore d'évaluation
- Devenir Cellule SexuelleDocument8 pagesDevenir Cellule SexuelleNookie StudiosPas encore d'évaluation
- Chap 5 FécondationDocument11 pagesChap 5 Fécondationabdoul djalil soumanaPas encore d'évaluation
- La Fecondation: Classe:3 C ProfesseurDocument6 pagesLa Fecondation: Classe:3 C Professeurepiphane diogoye mbenguePas encore d'évaluation
- Cours - SVT - La Procréation - Bac Math (2019-2020) MR Salhi MohamedDocument15 pagesCours - SVT - La Procréation - Bac Math (2019-2020) MR Salhi MohamedSassi LassaadPas encore d'évaluation
- Fécondation Et Intéraction Gamétique M. Anav PDFDocument45 pagesFécondation Et Intéraction Gamétique M. Anav PDFPapy PowerPas encore d'évaluation
- FécondationDocument10 pagesFécondationhauswirthPas encore d'évaluation
- 612756d3d8477la Reproduction Chez Les Mammiferes SVT TerminaleDocument6 pages612756d3d8477la Reproduction Chez Les Mammiferes SVT TerminaleAmour ConfiancePas encore d'évaluation
- FecondtionDocument6 pagesFecondtionRania ChitourPas encore d'évaluation
- UE2-Embryo-3-Fécondation (Final)Document22 pagesUE2-Embryo-3-Fécondation (Final)sun-nee-chan9Pas encore d'évaluation
- UTF-8 B bGEgZu - vWNvbmRhdGlvbiDvv71jb2xlIGRlIHNhbnTvv70 - UTF-8 B IFtNb2RlIGRlICBjb21wYXRpYmlsaXTvv71dLnBkZg - 1Document27 pagesUTF-8 B bGEgZu - vWNvbmRhdGlvbiDvv71jb2xlIGRlIHNhbnTvv70 - UTF-8 B IFtNb2RlIGRlICBjb21wYXRpYmlsaXTvv71dLnBkZg - 1Nour 1994Pas encore d'évaluation
- 2-Fécondation Contraception AMPDocument49 pages2-Fécondation Contraception AMPAy ManPas encore d'évaluation
- RESUME. FecondationDocument8 pagesRESUME. FecondationN'guessan Etienne BonnyPas encore d'évaluation
- Chapitre 3 Procreation Resume Du CoursDocument7 pagesChapitre 3 Procreation Resume Du Coursmaram mellekhPas encore d'évaluation
- 6-La FécondationDocument2 pages6-La FécondationahlemmPas encore d'évaluation
- Le Développement Embryonnaire Des AmphibiensDocument12 pagesLe Développement Embryonnaire Des AmphibiensKew27NaPas encore d'évaluation
- Gamatogenese Et FecondationDocument47 pagesGamatogenese Et FecondationAmïn Fïlalï100% (1)
- FécondationDocument47 pagesFécondationEric Charles HadjiPas encore d'évaluation
- Fécondation Fusion Des NoyauxDocument3 pagesFécondation Fusion Des NoyauxvivianogabrielpPas encore d'évaluation
- ExtraitDocument8 pagesExtraitremi gagbePas encore d'évaluation
- Chapitre 3 La Fécondation Alouani 2017Document7 pagesChapitre 3 La Fécondation Alouani 2017chahinda lakroutPas encore d'évaluation
- EXPOSERDocument38 pagesEXPOSERCheick SANOUPas encore d'évaluation
- Master 2 (Chap.1)Document6 pagesMaster 2 (Chap.1)Ažį ŽaPas encore d'évaluation
- Correction de La Série Fécondation Bac MathsDocument2 pagesCorrection de La Série Fécondation Bac MathsaretttbassPas encore d'évaluation
- Chapitre II SpermatogenèseDocument6 pagesChapitre II SpermatogenèseDel HadPas encore d'évaluation
- Embryologie Humaine GénéraleDocument13 pagesEmbryologie Humaine GénéraleHugues Roodly JeannitePas encore d'évaluation
- 1ère Semaine OkDocument56 pages1ère Semaine OkAÏCHATA KonéPas encore d'évaluation
- 4 FecondationDocument18 pages4 FecondationMACON824Pas encore d'évaluation
- Acc - FecondationDocument6 pagesAcc - Fecondationchahdsouf086Pas encore d'évaluation
- SVT TD - L8 - Le Devenir Des Cellules Sexuelles Chez Les MammifèresDocument6 pagesSVT TD - L8 - Le Devenir Des Cellules Sexuelles Chez Les MammifèresNiangPas encore d'évaluation
- Résumé - GamétogénèseDocument3 pagesRésumé - GamétogénèseRafaPas encore d'évaluation
- 2 GamétogenèseDocument12 pages2 GamétogenèseTCHETAN AmourPas encore d'évaluation
- Le Cycle Cellulaire Mon Cours PolycopeDocument26 pagesLe Cycle Cellulaire Mon Cours PolycopemohamedPas encore d'évaluation
- TDN 6 Corrigé Type PDFDocument3 pagesTDN 6 Corrigé Type PDFDiack Modou100% (1)
- La FecondationDocument53 pagesLa FecondationYaya Traoré100% (1)
- Procréation: Les Grands Articles d'UniversalisD'EverandProcréation: Les Grands Articles d'UniversalisPas encore d'évaluation
- Fonctionnement Global Des ÉcosystèmesDocument13 pagesFonctionnement Global Des ÉcosystèmesOussama Bouchetit100% (1)
- Fiche 04 Ac AntinucleaireDocument7 pagesFiche 04 Ac AntinucleaireNabil BahouraPas encore d'évaluation
- Correction TP6Document3 pagesCorrection TP6BskPas encore d'évaluation
- Généralités Sur Les BactériesDocument32 pagesGénéralités Sur Les Bactériesjuste ben johnsonPas encore d'évaluation
- Révision Sur Les Relations Alimentaires Avec Les PhotosDocument5 pagesRévision Sur Les Relations Alimentaires Avec Les PhotosIncandescent MostafaPas encore d'évaluation
- 201 Ex 197 Transplantation DorganeDocument5 pages201 Ex 197 Transplantation DorganeHmaied BaratliPas encore d'évaluation
- Guide Antibiotherapie Nac 2017Document116 pagesGuide Antibiotherapie Nac 2017Noriane RhouyPas encore d'évaluation
- Cours 01 G Enzy 1Document8 pagesCours 01 G Enzy 1sarraboulanouar79Pas encore d'évaluation
- Anatomie Et Physiologie Du Système Nerveux 2-1Document68 pagesAnatomie Et Physiologie Du Système Nerveux 2-1Abderrahmane DRISSIPas encore d'évaluation
- Atelier Interprétation ATBG PERPIGNAN CL AP 190919Document100 pagesAtelier Interprétation ATBG PERPIGNAN CL AP 190919Yomna Ben Lamine100% (1)
- Cours 6Document49 pagesCours 6Marwa BonitaPas encore d'évaluation
- 4-Les Milieux de Culture Utilises en Mycologie - 240312 - 113749Document59 pages4-Les Milieux de Culture Utilises en Mycologie - 240312 - 113749SANEK : Société d'Analyses et d'Essais KairouanPas encore d'évaluation
- Cours 5 Synthese Proteines EtudiantsDocument9 pagesCours 5 Synthese Proteines EtudiantsCita CitaPas encore d'évaluation
- La Nature de L'information GenetiqueDocument6 pagesLa Nature de L'information Genetiqueyassirfaddali2008Pas encore d'évaluation
- Candidoses VulvovaginalesDocument46 pagesCandidoses Vulvovaginaleswooden latexPas encore d'évaluation
- C4-INTRODUCTION À La MYCOLOGIEDocument71 pagesC4-INTRODUCTION À La MYCOLOGIEtabrose2607Pas encore d'évaluation
- Section A: Emploi Du Temps L3 Biologie Et Physiologie VégétaleDocument2 pagesSection A: Emploi Du Temps L3 Biologie Et Physiologie VégétaleZouheyrPas encore d'évaluation
- Ecologie GeneraleDocument126 pagesEcologie GeneraleInfamara Mané100% (2)
- Expose Sur Les OGMDocument10 pagesExpose Sur Les OGMReggad Elhadj0% (1)
- Le Consolateur - Chico XavierDocument410 pagesLe Consolateur - Chico XavierPlanète Révélations67% (6)
- Concours Blanc 6 BiologieDocument8 pagesConcours Blanc 6 Biologiejunior gatientPas encore d'évaluation
- PhytoestrogenesDocument1 pagePhytoestrogenesGuillaume michiganPas encore d'évaluation
- Corrigé DC 1 3SC 2022Document2 pagesCorrigé DC 1 3SC 2022iyeszoghlami206Pas encore d'évaluation
- Les Fonction de La Nutrition Chez L - HommeDocument2 pagesLes Fonction de La Nutrition Chez L - Hommeone asslanPas encore d'évaluation
- RTU GlucoseDocument3 pagesRTU Glucoseroumayssaa.yPas encore d'évaluation
- Présentation1 MicrobeDocument2 pagesPrésentation1 MicrobePrasanna Daniel100% (5)