Vous aimerez peut-être aussi
- ImmunoDocument5 pagesImmunoAmadou CISSEPas encore d'évaluation
- 97 1bDocument8 pages97 1bhizballah27Pas encore d'évaluation
- Suite Cours D - Immunologie PDFDocument61 pagesSuite Cours D - Immunologie PDFfafi Ali100% (1)
- 1.introduction À L'immunologie - Résumé HS PDFDocument3 pages1.introduction À L'immunologie - Résumé HS PDFbib bib100% (1)
- Le Systeme ImmunitaireDocument32 pagesLe Systeme ImmunitaireIlyes MraidiaPas encore d'évaluation
- Cours 1Document71 pagesCours 1midoan100% (1)
- ImmunoDocument29 pagesImmunoIkram DahmaniPas encore d'évaluation
- Concepts Fondamentaux en ImmunologieDocument42 pagesConcepts Fondamentaux en ImmunologieHajar ChehmatPas encore d'évaluation
- Cours ImmunoDocument29 pagesCours ImmunoChristian NgalaPas encore d'évaluation
- Système ImmunitaireDocument88 pagesSystème Immunitairehaniakamah2021Pas encore d'évaluation
- L'Immunité SpécifiqueDocument12 pagesL'Immunité Spécifiqueraslenbouallegue12345Pas encore d'évaluation
- Cours Chap 5Document7 pagesCours Chap 5Thevoice KidsPas encore d'évaluation
- AZ ImmunologieDocument33 pagesAZ ImmunologieFarahPas encore d'évaluation
- Notions D'immunologieDocument12 pagesNotions D'immunologieLucia MiguelPas encore d'évaluation
- CMH TB1920Document24 pagesCMH TB1920UrBehavior IsSo UghPas encore d'évaluation
- CMH Cours 4e A Pharm. Constantine 1Document10 pagesCMH Cours 4e A Pharm. Constantine 16hf7nzqkjbPas encore d'évaluation
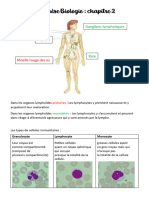
- Synthèse Biologie chp2Document6 pagesSynthèse Biologie chp2clara.plv22Pas encore d'évaluation
- Chapitre 6 - Le CMH - Etudiants PDFDocument3 pagesChapitre 6 - Le CMH - Etudiants PDFLennaPas encore d'évaluation
- Immunologie - Cahier PRDocument64 pagesImmunologie - Cahier PRJulie LEMOINEPas encore d'évaluation
- Y3 - Immunologie - All Q103Document19 pagesY3 - Immunologie - All Q103Narath LongPas encore d'évaluation
- 1 Er Cours Immuno Appl BS 202 PDFDocument10 pages1 Er Cours Immuno Appl BS 202 PDFHasna BentPas encore d'évaluation
- 0806 Reanimation Vol17 N4 p407 - 411Document5 pages0806 Reanimation Vol17 N4 p407 - 411FaresPas encore d'évaluation
- Les Réponses Immunitaires DR MILOUDIDocument13 pagesLes Réponses Immunitaires DR MILOUDIFarah B. BtoushPas encore d'évaluation
- Chapitre II Organes Et Cellules de L Immunit PDF PDFDocument22 pagesChapitre II Organes Et Cellules de L Immunit PDF PDFسارة عبيديPas encore d'évaluation
- IntroDocument61 pagesIntroGHERMI .MPas encore d'évaluation
- 7.implications Du Syst Hla en MédecineDocument14 pages7.implications Du Syst Hla en MédecineHa NaàPas encore d'évaluation
- Immunologie 1Document52 pagesImmunologie 1Mustafa KouzritePas encore d'évaluation
- Immuno3an-Organes Tissus Lymphoides BouabDocument13 pagesImmuno3an-Organes Tissus Lymphoides BouabSeif SeifoPas encore d'évaluation
- Introduction Générale À LimmunologieDocument67 pagesIntroduction Générale À LimmunologieNabil holmes100% (1)
- 2eme Biotech 19-20 Immunologie Générale AllouniDocument67 pages2eme Biotech 19-20 Immunologie Générale AllouniMaï Ssa100% (1)
- Immunité Inné PDFDocument38 pagesImmunité Inné PDFMou The ScientistPas encore d'évaluation
- Présentation de Lantigène L3 2019-2020 MMDocument41 pagesPrésentation de Lantigène L3 2019-2020 MMcheikhahmadinejadtidianediaPas encore d'évaluation
- UE2.2S1 DVD18 SystemeimmunitaireparleDrBoucrault Supportdecours 2Document57 pagesUE2.2S1 DVD18 SystemeimmunitaireparleDrBoucrault Supportdecours 2selmabendidi13Pas encore d'évaluation
- Fiche IntroDocument3 pagesFiche IntrorequiemforathrowawayPas encore d'évaluation
- Cours ImmunologieDocument285 pagesCours ImmunologiedouinienPas encore d'évaluation
- Déroulement de La Réponse Immunitaire SpécifiqueDocument6 pagesDéroulement de La Réponse Immunitaire SpécifiqueDalili SVTPas encore d'évaluation
- Chapitre IV, Les Mécanismes de L'immunité-1Document14 pagesChapitre IV, Les Mécanismes de L'immunité-1Balayira BakaryPas encore d'évaluation
- preparation علم المناعة 2 biofDocument7 pagespreparation علم المناعة 2 biofKotbi MohamedPas encore d'évaluation
- Immuno 3eme Annee EMD 1 (2015-2016)Document5 pagesImmuno 3eme Annee EMD 1 (2015-2016)Walid Bagh100% (2)
- Solution Des Exercices Le Système ImmunitaireDocument15 pagesSolution Des Exercices Le Système ImmunitairendiayePas encore d'évaluation
- Cour4 Expression D'antigènes, Marqueurs de Virulence Et de Récepteurs CellulairesDocument6 pagesCour4 Expression D'antigènes, Marqueurs de Virulence Et de Récepteurs CellulairesJoie De VivrePas encore d'évaluation
- Système immunitaire: Les Grands Articles d'UniversalisD'EverandSystème immunitaire: Les Grands Articles d'UniversalisPas encore d'évaluation
- ImmunologieDocument13 pagesImmunologiesamarasamounaPas encore d'évaluation
- Ronéos HématologieDocument123 pagesRonéos HématologieMohamed-Amine SADKIPas encore d'évaluation
- Cours Chapitre IIDocument7 pagesCours Chapitre IIErwan GuioniePas encore d'évaluation
- Bilan Chapitre 3 L ' Immunite AdaptativeDocument6 pagesBilan Chapitre 3 L ' Immunite Adaptativeahmedrayanelali23Pas encore d'évaluation
- المستندDocument4 pagesالمستندlakhfksnufPas encore d'évaluation
- 2 AntigenesDocument6 pages2 Antigenesibrahima kanePas encore d'évaluation
- Immunologie IDocument107 pagesImmunologie Iمصطفى الورديغيPas encore d'évaluation
- 05 - CMH Résidents 2015Document93 pages05 - CMH Résidents 2015liliwoody986Pas encore d'évaluation
- FR - Semaine 1 - Final PDFDocument43 pagesFR - Semaine 1 - Final PDFChristian GaudinPas encore d'évaluation
- Essentiel11 Immunite AdaptativeDocument2 pagesEssentiel11 Immunite Adaptativejad mamouPas encore d'évaluation
- Chapitre 1 - Introduction À L - Immunologie - ElumensDocument9 pagesChapitre 1 - Introduction À L - Immunologie - ElumenstructresdouteuxPas encore d'évaluation
- Immunologie. CMHDocument8 pagesImmunologie. CMHËl kíng čhrístíän MükänïrwäPas encore d'évaluation
- JjiiijjDocument31 pagesJjiiijjFaith CyrilPas encore d'évaluation
- Les Écoles de l'APBI: BiologieDocument12 pagesLes Écoles de l'APBI: BiologieHackbx2 Mafia netPas encore d'évaluation
- Chap 1 RÃcepteurs Et Les Cellules de Limmunitã InnÃe PDFDocument17 pagesChap 1 RÃcepteurs Et Les Cellules de Limmunitã InnÃe PDFikram BencherifPas encore d'évaluation
- 1 - Introduction À L'immunologieDocument4 pages1 - Introduction À L'immunologieOsama O-bPas encore d'évaluation
- L'Immunologie - SVT 1èrea Tome2Document7 pagesL'Immunologie - SVT 1èrea Tome2Guy TangaPas encore d'évaluation