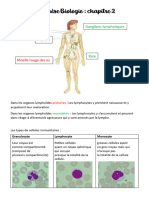
Vous aimerez peut-être aussi
- Système immunitaire: Les Grands Articles d'UniversalisD'EverandSystème immunitaire: Les Grands Articles d'UniversalisPas encore d'évaluation
- Cours 1Document71 pagesCours 1midoan100% (1)
- Resumé Du Cours de L'immunitéDocument3 pagesResumé Du Cours de L'immunitéMomme Ould Deyahi Achrif100% (8)
- Immunologie 1Document52 pagesImmunologie 1Mustafa KouzritePas encore d'évaluation
- RB. Guide Pour Le Choix Des Désinfectant PDFDocument110 pagesRB. Guide Pour Le Choix Des Désinfectant PDFfourat.zarkounaPas encore d'évaluation
- Les StreptocoquesDocument8 pagesLes Streptocoquesismail bd100% (1)
- Chapitre 13 L'immunité Adaptative, Prolongement de L'immunité InnéeDocument12 pagesChapitre 13 L'immunité Adaptative, Prolongement de L'immunité InnéeSparrowPas encore d'évaluation
- Tuberculose OsseuseDocument67 pagesTuberculose OsseuseImene SDPas encore d'évaluation
- 1.introduction À L'immunologie - Résumé HS PDFDocument3 pages1.introduction À L'immunologie - Résumé HS PDFbib bib100% (1)
- Le Systeme ImmunitaireDocument32 pagesLe Systeme ImmunitaireIlyes MraidiaPas encore d'évaluation
- Déroulement de La Réponse Immunitaire Spécifique-ConvertiDocument20 pagesDéroulement de La Réponse Immunitaire Spécifique-ConvertiHk Eh100% (2)
- Immunite CoursDocument14 pagesImmunite CoursJulien GhandourPas encore d'évaluation
- ImmunoDocument29 pagesImmunoIkram DahmaniPas encore d'évaluation
- Déroulement de La Réponse Immunitaire SpécifiqueDocument6 pagesDéroulement de La Réponse Immunitaire SpécifiqueDalili SVTPas encore d'évaluation
- 2.réaction. Précipitation Et AgglutinationDocument9 pages2.réaction. Précipitation Et AgglutinationRomi Atoui100% (1)
- Bilan Chapitre 3 L ' Immunite AdaptativeDocument6 pagesBilan Chapitre 3 L ' Immunite Adaptativeahmedrayanelali23Pas encore d'évaluation
- Chap 15Document3 pagesChap 15Frédéric JanvierPas encore d'évaluation
- Cours Chap 5Document7 pagesCours Chap 5Thevoice KidsPas encore d'évaluation
- Essentiel11 Immunite AdaptativeDocument2 pagesEssentiel11 Immunite Adaptativejad mamouPas encore d'évaluation
- Cours Médiation HumoraleDocument12 pagesCours Médiation Humoraleel adab yesminePas encore d'évaluation
- Immuno 2Document28 pagesImmuno 2Ameen MaoPas encore d'évaluation
- Ts Th3a Chap2 Cours Reaction Adaptative-2Document12 pagesTs Th3a Chap2 Cours Reaction Adaptative-2Jibril Berrando BerrPas encore d'évaluation
- preparation علم المناعة 2 biofDocument7 pagespreparation علم المناعة 2 biofKotbi MohamedPas encore d'évaluation
- Solution Des Exercices Le Système ImmunitaireDocument15 pagesSolution Des Exercices Le Système ImmunitairendiayePas encore d'évaluation
- Synthèse Biologie chp2Document6 pagesSynthèse Biologie chp2clara.plv22Pas encore d'évaluation
- ImmunologieDocument33 pagesImmunologieChrisberle ZePas encore d'évaluation
- L'Immunité SpécifiqueDocument12 pagesL'Immunité Spécifiqueraslenbouallegue12345Pas encore d'évaluation
- ImmunologieDocument10 pagesImmunologieLilian Saint-LégerPas encore d'évaluation
- GP26Document1 pageGP26vchhuhPas encore d'évaluation
- Activite-Origine Anticorps CorrigeDocument5 pagesActivite-Origine Anticorps CorrigeThevoice KidsPas encore d'évaluation
- Support de Cours R Ponse Immunitaire Humorale Et CellulaireDocument12 pagesSupport de Cours R Ponse Immunitaire Humorale Et CellulaireBOURRAKH HAMZAPas encore d'évaluation
- Magazine Le Deroulement de La Reponse ImmunitaireDocument19 pagesMagazine Le Deroulement de La Reponse ImmunitaireMohamed Helmi Chebbi100% (1)
- ImmunoDocument5 pagesImmunoAmadou CISSEPas encore d'évaluation
- Thème 3 Chapitre 3Document27 pagesThème 3 Chapitre 3TIMCCPas encore d'évaluation
- Mécanisme de Défense SpécifiqueDocument6 pagesMécanisme de Défense SpécifiqueJoseph OnanaPas encore d'évaluation
- La Reconnaissance Du Soi Et Du Non SoiDocument6 pagesLa Reconnaissance Du Soi Et Du Non SoiRonan PerennesPas encore d'évaluation
- Chapitre 2-7Document3 pagesChapitre 2-7hadidsciencesPas encore d'évaluation
- Immunologie 2Document110 pagesImmunologie 2Ayoub tahiriPas encore d'évaluation
- Immuon 000Document2 pagesImmuon 000bouhamlamanarchemselyakinePas encore d'évaluation
- Fax Immunologie ReduitDocument2 pagesFax Immunologie Reduitissaalimahamat1998Pas encore d'évaluation
- Cours ImmunologieDocument285 pagesCours ImmunologiedouinienPas encore d'évaluation
- L Immunite Adaptative Cours 1Document10 pagesL Immunite Adaptative Cours 1Christian abahPas encore d'évaluation
- Leçon2 ÉlèveDocument4 pagesLeçon2 ÉlèveMarie PoettiPas encore d'évaluation
- Chapitre 1 - Introduction À L - Immunologie - ElumensDocument9 pagesChapitre 1 - Introduction À L - Immunologie - ElumenstructresdouteuxPas encore d'évaluation
- Theme 3 Chapitre 16 3 Et 4Document2 pagesTheme 3 Chapitre 16 3 Et 4louislucasbaptistaPas encore d'évaluation
- Corrigé La Maturation Du Systeme Immunitaire en 3 Points.Document4 pagesCorrigé La Maturation Du Systeme Immunitaire en 3 Points.kilm. gujo.Pas encore d'évaluation
- SN16-TE-06-19-Chapitre 2Document16 pagesSN16-TE-06-19-Chapitre 2claudeulrichomgbatabi237Pas encore d'évaluation
- Immunologie - Cahier PRDocument64 pagesImmunologie - Cahier PRJulie LEMOINEPas encore d'évaluation
- Mécanisme de La Réponse ImmunitaireDocument12 pagesMécanisme de La Réponse ImmunitaireKurtisgangue18Pas encore d'évaluation
- Suite Cours D - Immunologie PDFDocument61 pagesSuite Cours D - Immunologie PDFfafi Ali100% (1)
- 3è TH 3 Le SI Cours P 5-6Document2 pages3è TH 3 Le SI Cours P 5-6Mohammed Yassir SAADANI HASSANIPas encore d'évaluation
- ImmunologieDocument28 pagesImmunologieImenePas encore d'évaluation
- Fiche IntroDocument3 pagesFiche IntrorequiemforathrowawayPas encore d'évaluation
- Immuno3an-Reponses Immunitaires2018benachourDocument9 pagesImmuno3an-Reponses Immunitaires2018benachourAmina MnPas encore d'évaluation
- Cours Immunité Adaptative PDFDocument6 pagesCours Immunité Adaptative PDFL kPas encore d'évaluation
- Terminale S Fiche Revision ImmunologieDocument4 pagesTerminale S Fiche Revision ImmunologieMiantsa Ramahaliarivo100% (1)
- ImmunologieDocument6 pagesImmunologiemalak wxPas encore d'évaluation
- Exposé Sur L'immunité, L'anticors, L'antigène, La Phagosytose Ect....Document36 pagesExposé Sur L'immunité, L'anticors, L'antigène, La Phagosytose Ect....OUSSAMA100% (1)
- B) Amplification ClonaleDocument3 pagesB) Amplification ClonaleOwen DormoisPas encore d'évaluation
- 97 1bDocument8 pages97 1bhizballah27Pas encore d'évaluation
- Cours IV Tolérance Immunitaire Et Auto ImmunitéDocument13 pagesCours IV Tolérance Immunitaire Et Auto ImmunitétinaPas encore d'évaluation
- Cours 3B Du 18 AvrilDocument6 pagesCours 3B Du 18 AvriladjehmiPas encore d'évaluation
- Cours Immunologie S5Document113 pagesCours Immunologie S5hamzaabzou154Pas encore d'évaluation
- 4-NK CellsDocument23 pages4-NK Cellssanaahlem0Pas encore d'évaluation
- Version 1Document23 pagesVersion 1hamache imenePas encore d'évaluation
- Génétique Et CorrectionDocument3 pagesGénétique Et CorrectionsafaelgarehPas encore d'évaluation
- Appretement Et Presentation de L'agDocument41 pagesAppretement Et Presentation de L'agAnonymous FGGLsjzWi100% (2)
- TD Génétique L1 - USTSDocument8 pagesTD Génétique L1 - USTSsimaedouaaron067Pas encore d'évaluation
- Lvrea 266 P 517Document6 pagesLvrea 266 P 517Samira LarouciPas encore d'évaluation
- Anti Biot I QuesDocument49 pagesAnti Biot I QuesRekia BenPas encore d'évaluation
- Transmission Des Caracteres 2019Document36 pagesTransmission Des Caracteres 2019Abdo AichaPas encore d'évaluation
- Cours IV Tolérance Immunitaire Et Auto ImmunitéDocument13 pagesCours IV Tolérance Immunitaire Et Auto ImmunitétinaPas encore d'évaluation
- Profil Epidemiologique Et de Resistance Des Bacteries Multi Resistantes Au Chu Hassan Ii de FèsDocument112 pagesProfil Epidemiologique Et de Resistance Des Bacteries Multi Resistantes Au Chu Hassan Ii de Fèsx-Abdelali Ait ouaichaPas encore d'évaluation
- CH 3 AbnaDocument11 pagesCH 3 AbnaRym SekiouPas encore d'évaluation
- TD 2 MbioDocument33 pagesTD 2 MbioliliroseplPas encore d'évaluation
- EOI FR2 Tema 2.1 Je Suis Malade Une Sante de FerDocument55 pagesEOI FR2 Tema 2.1 Je Suis Malade Une Sante de FerAdrián Alarcón OrtizPas encore d'évaluation
- SVT Num 3Document1 pageSVT Num 3Simohamed Gassa GassaPas encore d'évaluation
- Obstétrique Des Petits AnimauxDocument24 pagesObstétrique Des Petits AnimauxCharlotte Taton100% (1)
- Chapitre 3Document5 pagesChapitre 3bouchakour meryemPas encore d'évaluation
- Contrôle Des Aliments VFDocument216 pagesContrôle Des Aliments VFmanalyoubi00Pas encore d'évaluation
- CourDocument8 pagesCourKlanzy MandPas encore d'évaluation
- Rév Gen 2020Document39 pagesRév Gen 2020Mohamed Helmi ChebbiPas encore d'évaluation
- Bactéries Des Plaies Infectées Suite À Morsure HumaineDocument3 pagesBactéries Des Plaies Infectées Suite À Morsure Humainesfendri17Pas encore d'évaluation
- Cours de Génomique, Master BH, S1, DR OKON, 2020-2021Document34 pagesCours de Génomique, Master BH, S1, DR OKON, 2020-2021KONAN ABO PHILEMON ARONOUPas encore d'évaluation
- CoronaDocument2 pagesCoronaHamidi FaizaPas encore d'évaluation
- Cours Microorganismes Et Santé Des PlantesDocument53 pagesCours Microorganismes Et Santé Des PlantesAbdessamad ImoulanPas encore d'évaluation
- Item 151 Infections Broncho Pulmonaires Communautaires de L Adulte Et de L EnfantDocument19 pagesItem 151 Infections Broncho Pulmonaires Communautaires de L Adulte Et de L EnfantcacapizonPas encore d'évaluation
- 12 2004 FR 2Document3 pages12 2004 FR 2Jay MenonPas encore d'évaluation