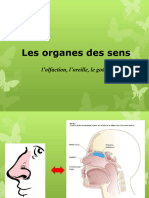
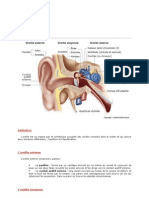
Vous aimerez peut-être aussi
- Hygiène de la voix et gymnastique des organes vocaux: Histoire de la musique depuis son origine jusqu'à nos joursD'EverandHygiène de la voix et gymnastique des organes vocaux: Histoire de la musique depuis son origine jusqu'à nos joursPas encore d'évaluation
- Anatomie Et Physiologie en ORLDocument77 pagesAnatomie Et Physiologie en ORLbenben31Pas encore d'évaluation
- Chapitre 11 - Les Systèmes MentalesDocument66 pagesChapitre 11 - Les Systèmes MentalesMouctarPas encore d'évaluation
- 5 Sens-53527Document14 pages5 Sens-53527Iuliana ErhanPas encore d'évaluation
- Entendre La MusiqueDocument3 pagesEntendre La MusiqueLaurène MangenotPas encore d'évaluation
- Expose de Medecine PhysiqueDocument8 pagesExpose de Medecine Physiqueamitthai MasengoPas encore d'évaluation
- Séance 1Document12 pagesSéance 1alexismoutierPas encore d'évaluation
- Anatomie Organes de SensDocument28 pagesAnatomie Organes de SensdjouadjonbelvigePas encore d'évaluation
- Cours de L' OlfactionDocument6 pagesCours de L' OlfactionDeaƭh SurgeơnPas encore d'évaluation
- Curs 3 UrecheaDocument28 pagesCurs 3 Urecheaqty9jgkpnzPas encore d'évaluation
- La PsychoacoustiqueDocument22 pagesLa PsychoacoustiqueHumzy100% (1)
- NEUROANAT2023 (Jude Thiry) (1) - Part-3Document71 pagesNEUROANAT2023 (Jude Thiry) (1) - Part-3EligarciadewitPas encore d'évaluation
- 5 SensDocument3 pages5 SensDrira AnisPas encore d'évaluation
- 5eme SéanceDocument35 pages5eme SéanceMohamed AlouanePas encore d'évaluation
- Anatomie Oreille Cours IPFTSDocument9 pagesAnatomie Oreille Cours IPFTSYassine BoukriPas encore d'évaluation
- 7 - Organe de L'olfactionDocument30 pages7 - Organe de L'olfactionAmmar ChaouiPas encore d'évaluation
- Organes Des SensDocument12 pagesOrganes Des SensEl Zizou MerenguePas encore d'évaluation
- 2.neurophysiologie de L'auditionDocument12 pages2.neurophysiologie de L'auditionboutefal imenePas encore d'évaluation
- Document Sans TitreDocument7 pagesDocument Sans TitresisiladnsPas encore d'évaluation
- Ch. 1. SyntheseDocument5 pagesCh. 1. Synthesenokiagaming1234Pas encore d'évaluation
- Ba 97 Aba 3Document2 pagesBa 97 Aba 3Aymane wahmanePas encore d'évaluation
- Voies Aériennes Supérieures (Cour)Document5 pagesVoies Aériennes Supérieures (Cour)Medjoudj SarahPas encore d'évaluation
- Le Nerf OlfactifDocument9 pagesLe Nerf OlfactifDanielle MeyoPas encore d'évaluation
- TP2 Oreille Interne Version Travail MaisonDocument2 pagesTP2 Oreille Interne Version Travail Maisonzorojuro3934Pas encore d'évaluation
- Chapitre I. Implants Cochléaires Et Stratégies de StimulationDocument20 pagesChapitre I. Implants Cochléaires Et Stratégies de StimulationelhoucinePas encore d'évaluation
- Fonctionnement de L'oreilleDocument14 pagesFonctionnement de L'oreilleSatch GuitarePas encore d'évaluation
- Fichier Produit 2315Document8 pagesFichier Produit 2315MaurelPas encore d'évaluation
- Cours Théme 4 1ére ESDocument5 pagesCours Théme 4 1ére ESSarah 18Pas encore d'évaluation
- 11 - Audition Dr. LassakeurDocument75 pages11 - Audition Dr. LassakeurAmmar ChaouiPas encore d'évaluation
- Cours 8. Les Organes Des SensDocument4 pagesCours 8. Les Organes Des SensmousalsaletturkiyaPas encore d'évaluation
- Les SensationsDocument26 pagesLes SensationsMOUSTAPHAPas encore d'évaluation
- Exposé - Tazi.kada - Reffas (3) CopieDocument8 pagesExposé - Tazi.kada - Reffas (3) Copietazimanel25Pas encore d'évaluation
- 50 Cours Ifsi Beziers 2Document40 pages50 Cours Ifsi Beziers 2DohaAitAntarPas encore d'évaluation
- L'oreille, L'ouie Et L'équilibreDocument2 pagesL'oreille, L'ouie Et L'équilibrenourimane22112009Pas encore d'évaluation
- M2-Neuro - Anatomie de L'audition Et de L'équilibrationDocument12 pagesM2-Neuro - Anatomie de L'audition Et de L'équilibrationThomas HussonPas encore d'évaluation
- Le Nez TexteDocument9 pagesLe Nez TexteAdognon Jaquet DeJésusPas encore d'évaluation
- Organe OlfactifDocument2 pagesOrgane Olfactifmalakzerizer2005Pas encore d'évaluation
- La Muqueuse Olfactive Et Les Bourgeons Du GoutDocument8 pagesLa Muqueuse Olfactive Et Les Bourgeons Du Goutjoudjojo.06Pas encore d'évaluation
- OlfactionDocument4 pagesOlfactionsoh harnaPas encore d'évaluation
- Histologie de L'appareil RespiratoireDocument7 pagesHistologie de L'appareil RespiratoireAhlam BsdPas encore d'évaluation
- C16. Organes de SensDocument10 pagesC16. Organes de SensTCHETAN AmourPas encore d'évaluation
- Histologie de L'oreille 2011-2012Document72 pagesHistologie de L'oreille 2011-2012Nada Affes89% (9)
- L'Appareil RespiratoireDocument10 pagesL'Appareil RespiratoireMark LandersPas encore d'évaluation
- Phenomene Objectif de L'audition (BI-chouiab-cherkeoui-ghait-benhessy)Document16 pagesPhenomene Objectif de L'audition (BI-chouiab-cherkeoui-ghait-benhessy)Hou Da GhaitPas encore d'évaluation
- Minimodule ORLDocument46 pagesMinimodule ORLibtihelantriPas encore d'évaluation
- Phésanimal Chirouf Akram Hicham 2 Sa Groupe 1Document36 pagesPhésanimal Chirouf Akram Hicham 2 Sa Groupe 1NOOBITO'sPas encore d'évaluation
- Organe de SensDocument10 pagesOrgane de Senswafa beyPas encore d'évaluation
- Physiologie Animale NemarDocument49 pagesPhysiologie Animale NemarOhomonPas encore d'évaluation
- 8 - Audition Equilibration 2021Document89 pages8 - Audition Equilibration 2021Ammar ChaouiPas encore d'évaluation
- Syst RespiDocument44 pagesSyst Respiantoineprevot1999Pas encore d'évaluation
- Transmission de L'onde SonoreDocument39 pagesTransmission de L'onde Sonoreadelmiringui2Pas encore d'évaluation
- DocumentDocument16 pagesDocumentAdriana FongangPas encore d'évaluation
- Entendre La MusiqueDocument2 pagesEntendre La Musiqueemadalkasmi2Pas encore d'évaluation
- Legende Oreille V2 TP Oreill Et AuditionDocument1 pageLegende Oreille V2 TP Oreill Et Auditionsoufireda25Pas encore d'évaluation
- AuditionDocument24 pagesAuditionPercheryPas encore d'évaluation
- Le Système RespiratoireDocument22 pagesLe Système RespiratoiretoufikPas encore d'évaluation
- 22 La Cochlée Membraneuse Et Voies CochléairesDocument5 pages22 La Cochlée Membraneuse Et Voies Cochléairesadelmiringui2Pas encore d'évaluation
- Anatomi de L'oreilleDocument2 pagesAnatomi de L'oreillemerymorenaaPas encore d'évaluation
- 2-Histologie de L'Appareil RespiratoireDocument61 pages2-Histologie de L'Appareil RespiratoireImene Imene100% (1)
- 1ère C Et D Ens SC SVT 30.04 8h.9h ABRAHAM PDFDocument4 pages1ère C Et D Ens SC SVT 30.04 8h.9h ABRAHAM PDFriad ouastiPas encore d'évaluation
- Neuroanatomie Funciones-LexicoDocument69 pagesNeuroanatomie Funciones-LexicoDaniela RiosPas encore d'évaluation
- Goleman, Daniel - L'Intelligence ÉmotionnelleDocument249 pagesGoleman, Daniel - L'Intelligence ÉmotionnelleAnonymous w1iCBlT100% (11)
- Médecine 13 Les Nerfs Craniens Thomas Robert 2021 08 02 OkDocument106 pagesMédecine 13 Les Nerfs Craniens Thomas Robert 2021 08 02 Okdidier ramatisPas encore d'évaluation
- L'odorat Et Ses TroublesDocument109 pagesL'odorat Et Ses TroublesNicholas JeffriesPas encore d'évaluation
- $atteintes Isolees Des Nerfs Craniens Old EMCDocument63 pages$atteintes Isolees Des Nerfs Craniens Old EMCEddy SaadPas encore d'évaluation
- 7 - Organe de L'olfactionDocument30 pages7 - Organe de L'olfactionAmmar ChaouiPas encore d'évaluation
- 1 L'Audition, L'odorat, Le Goût, Le ToucherDocument24 pages1 L'Audition, L'odorat, Le Goût, Le ToucherYohanPas encore d'évaluation
- 2-Résumé Lydia Tadjadit Part2 PDFDocument42 pages2-Résumé Lydia Tadjadit Part2 PDFSaibi100% (1)
- Le Système Sensoriel COURSDocument95 pagesLe Système Sensoriel COURSRebecca MoulackaPas encore d'évaluation
- Memoire GuyauxDocument134 pagesMemoire GuyauxViviane TjiovasPas encore d'évaluation