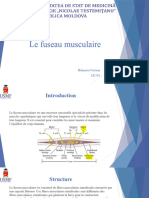
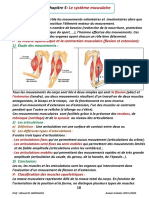
Vous aimerez peut-être aussi
- Cours Système NerveuxDocument25 pagesCours Système NerveuxAblaye100% (1)
- Comptabilité Des CliniquesDocument47 pagesComptabilité Des CliniquesAnonymous kAVA6ALXNPas encore d'évaluation
- Système Nerveux 1Document21 pagesSystème Nerveux 1Ranime Mekersi100% (1)
- Met S 23 Sy PDocument4 pagesMet S 23 Sy Pkhocine100% (1)
- Muscles: Les Grands Articles d'UniversalisD'EverandMuscles: Les Grands Articles d'UniversalisPas encore d'évaluation
- 2-La Hierarchie AngéliqueDocument6 pages2-La Hierarchie Angéliquemichel fayettetchibinda100% (2)
- Robe Salopette CatherineDocument57 pagesRobe Salopette CatherineAude EPIARD100% (1)
- Les Vrais Secrets de La Magie NoireDocument113 pagesLes Vrais Secrets de La Magie NoireKawtar Gloom GOthside ൃ100% (1)
- 3718 v2 Planchers Nervurs Poutrelles Prfa Associes Du Bton Coul en OeuvreDocument98 pages3718 v2 Planchers Nervurs Poutrelles Prfa Associes Du Bton Coul en OeuvreZizouk FeresPas encore d'évaluation
- Physiologie Musculaire 2020Document47 pagesPhysiologie Musculaire 2020drahlemtrifPas encore d'évaluation
- II Les Réflexes ProprioceptifsDocument31 pagesII Les Réflexes ProprioceptifsHamza Abu Humaid100% (2)
- Physiologie Neuro-MuscuclaireDocument53 pagesPhysiologie Neuro-MuscuclaireMampionona RakotonandrasanaPas encore d'évaluation
- Pied Au PlancherDocument2 pagesPied Au PlancherKenza BenmokhtarPas encore d'évaluation
- NeurophysiologieDocument13 pagesNeurophysiologieCath Lemeunier100% (1)
- La Contraction MusculaireDocument5 pagesLa Contraction MusculairekharratPas encore d'évaluation
- Lantschoot, Arnold Van. Un Texte Palimpseste de Vat. Copte 65, Muséon 60 (1947), 261-268Document8 pagesLantschoot, Arnold Van. Un Texte Palimpseste de Vat. Copte 65, Muséon 60 (1947), 261-268yadatanPas encore d'évaluation
- ContractionDocument13 pagesContractionPY00Pas encore d'évaluation
- Cours Myologie PsychomotDocument7 pagesCours Myologie PsychomotRazenAlgPas encore d'évaluation
- P Motricité (C. Quairiaux) - 26.03.21Document5 pagesP Motricité (C. Quairiaux) - 26.03.21nasten16fhPas encore d'évaluation
- Neuroanatomie FonctionnelleDocument14 pagesNeuroanatomie FonctionnelleWilliams SantimePas encore d'évaluation
- l3 Neurophysio Chap4Document16 pagesl3 Neurophysio Chap4nobodyPas encore d'évaluation
- Tonus MotriciteDocument15 pagesTonus MotricitecharbelhannaPas encore d'évaluation
- Le Muscle SquelletiqueDocument6 pagesLe Muscle SquelletiquefgouzikPas encore d'évaluation
- SN06TEWB0522 Retenir-PartieIDocument10 pagesSN06TEWB0522 Retenir-PartieIremandasarahPas encore d'évaluation
- SVT - Fiche 14 - RéflexesDocument8 pagesSVT - Fiche 14 - RéflexesClemence BrasseletPas encore d'évaluation
- Le Systeme Musculaire Serie d Exercices Word 1Document11 pagesLe Systeme Musculaire Serie d Exercices Word 1zinebsemlali2009Pas encore d'évaluation
- Le Système Musculaire_015620Document6 pagesLe Système Musculaire_015620مصعب صارديPas encore d'évaluation
- Le Fonctionnement Du Systeme NerveuxDocument11 pagesLe Fonctionnement Du Systeme NerveuxSadi KoukouPas encore d'évaluation
- Muscle Et Physiologie MusculaireDocument20 pagesMuscle Et Physiologie MusculaireBoubker PouperPas encore d'évaluation
- La Fonction Musculaire Te Le MouvementDocument54 pagesLa Fonction Musculaire Te Le Mouvementmosbahhafida18Pas encore d'évaluation
- Cours Mucle RénovéDocument11 pagesCours Mucle Rénovémohamed.aziz.amri.11Pas encore d'évaluation
- SVT Le Reflexe Myotatique PDFDocument7 pagesSVT Le Reflexe Myotatique PDFAnonymous TrrvM8Pas encore d'évaluation
- Système Musculaire 2020Document22 pagesSystème Musculaire 2020Sarah OuajnatiPas encore d'évaluation
- Physiologie de L'appareil Locomoteur-Le Système MusculaireDocument6 pagesPhysiologie de L'appareil Locomoteur-Le Système MusculaireEmy DesreumauxPas encore d'évaluation
- Système MusculaireDocument16 pagesSystème MusculaireMustapha BriachePas encore d'évaluation
- TD Reflexe MyotatiqueDocument4 pagesTD Reflexe MyotatiquelemiaPas encore d'évaluation
- Notes de Cours - La Motricité PériphériqueDocument8 pagesNotes de Cours - La Motricité PériphériqueanaischapellzPas encore d'évaluation
- Le fuseau neuromusculaire-1Document10 pagesLe fuseau neuromusculaire-1cristinarotaras2Pas encore d'évaluation
- MuscleDocument15 pagesMuscleMaximePas encore d'évaluation
- Le fuseau neuromusculaire-1Document10 pagesLe fuseau neuromusculaire-1cristinarotaras2Pas encore d'évaluation
- Le Syst Me MusculaireDocument27 pagesLe Syst Me MusculaireSarah Dos SantosPas encore d'évaluation
- Système MusculaireDocument7 pagesSystème MusculaireAmine WlPas encore d'évaluation
- Controle MedullaireDocument16 pagesControle MedullaireAminou MounirouPas encore d'évaluation
- TSpe T3B1 CoursDocument5 pagesTSpe T3B1 CoursFranckyscocom Gokpan Enfant BeniPas encore d'évaluation
- 7 - Les Tissus MusculairesDocument6 pages7 - Les Tissus MusculairesHayfa Ben taherPas encore d'évaluation
- Chapitre 3A Cours Sur Cerveau Et Mouvement VolontaireDocument3 pagesChapitre 3A Cours Sur Cerveau Et Mouvement Volontairealice.maranberPas encore d'évaluation
- FetchDocument13 pagesFetchAhmed RekikPas encore d'évaluation
- FA1 Correction-2Document9 pagesFA1 Correction-2Emmanuel MaxPas encore d'évaluation
- Contracté - Relâché, Encore Une Fois. - François RosenzweigDocument102 pagesContracté - Relâché, Encore Une Fois. - François RosenzweigOlivia ManceauPas encore d'évaluation
- SN4 Controle MoteurDocument5 pagesSN4 Controle Moteurhoussein zmerliPas encore d'évaluation
- Système Nerveux AutonomeDocument21 pagesSystème Nerveux AutonomeArtichaudPas encore d'évaluation
- La MyologieDocument6 pagesLa MyologieBilly Bel Vocale MesidorPas encore d'évaluation
- SNC Et SNVDocument3 pagesSNC Et SNVlenoirmccPas encore d'évaluation
- Chapitre 5:: Le Système MusculaireDocument5 pagesChapitre 5:: Le Système MusculaireRAYANE YTPas encore d'évaluation
- Chap 1 SYSTEME NERVEUX GENERALITESDocument7 pagesChap 1 SYSTEME NERVEUX GENERALITESAnani Jacques YaoPas encore d'évaluation
- Le-Systeme-Musculaire 2022 PDocument4 pagesLe-Systeme-Musculaire 2022 Pmarouane elhimaniPas encore d'évaluation
- Les Tissus MusculairesDocument14 pagesLes Tissus Musculairesariane.vasseur29Pas encore d'évaluation
- Le Système Musculaire 3ACDocument5 pagesLe Système Musculaire 3AC1mf2p100% (1)
- 2 - Physiologie de La MotricitéDocument115 pages2 - Physiologie de La MotricitéBouchafra AbderahimPas encore d'évaluation
- Cours 6. Le Systeme NerveuxDocument10 pagesCours 6. Le Systeme Nerveuxkado01nathaliaPas encore d'évaluation
- Tissu Musculaire MeksemDocument45 pagesTissu Musculaire MeksemNo Comment.Pas encore d'évaluation
- Chapitre Iv Les Cellules ExcitablesDocument77 pagesChapitre Iv Les Cellules ExcitablesSARA IBRAHIMIPas encore d'évaluation
- Neuro2 DR AlaouiDocument61 pagesNeuro2 DR Alaouiyousra.errebjaPas encore d'évaluation
- Neurophysiologie(1)Document15 pagesNeurophysiologie(1)aitbaali.oriflamePas encore d'évaluation
- La Moelle Épinière, Centre RéflexeDocument6 pagesLa Moelle Épinière, Centre RéflexeSaken667Pas encore d'évaluation
- Physio SNDocument51 pagesPhysio SNNo Comment.Pas encore d'évaluation
- Support N° 8Document7 pagesSupport N° 8Ossete BricePas encore d'évaluation
- Sys867 Sujets SP É Ciaux I: Traitement Des Signaux Biom É Dicaux É LectromyographieDocument56 pagesSys867 Sujets SP É Ciaux I: Traitement Des Signaux Biom É Dicaux É LectromyographieBelaliaPas encore d'évaluation
- 17M110hhhhhh PDFDocument101 pages17M110hhhhhh PDFilham haniniPas encore d'évaluation
- EchographieDocument10 pagesEchographieImaa Ha NnePas encore d'évaluation
- LibertéDocument19 pagesLibertételahamidPas encore d'évaluation
- Mécanique Des RochesDocument41 pagesMécanique Des RochesIssam ZougariPas encore d'évaluation
- Chapitre5 DHCPDocument17 pagesChapitre5 DHCPjeanPas encore d'évaluation
- Corro Philo Valesse 1iere - PDFDocument114 pagesCorro Philo Valesse 1iere - PDFnayoussayacoubou17Pas encore d'évaluation
- Regarder Le Tresor Du Petit Nicolas Entier VFDocument4 pagesRegarder Le Tresor Du Petit Nicolas Entier VFxzcvxzPas encore d'évaluation
- Rapport Cable TechDocument17 pagesRapport Cable TechMallouki Med100% (1)
- SubnettingDocument19 pagesSubnettingtunisianouPas encore d'évaluation
- Yadh Ben AchourDocument26 pagesYadh Ben AchourIsmail SadaouiPas encore d'évaluation
- Syl M2 Inf-DcDocument28 pagesSyl M2 Inf-DcChihebeddine AmmarPas encore d'évaluation
- Chimie GeneraleDocument5 pagesChimie Generaletcheva jokhanan TiambiPas encore d'évaluation
- Manuel D'utilisation Gamme Murale Ar5000 1.1 Samsung 2014Document57 pagesManuel D'utilisation Gamme Murale Ar5000 1.1 Samsung 2014ddrumeaPas encore d'évaluation
- TEXTE N°1: La Jeune AcrobateDocument5 pagesTEXTE N°1: La Jeune Acrobatelouaslama0106Pas encore d'évaluation
- Repercussion Islam Ames Hamad1Document15 pagesRepercussion Islam Ames Hamad1saydou bokotaPas encore d'évaluation
- Niveaux Langue Registres Exercices PDFDocument4 pagesNiveaux Langue Registres Exercices PDFSadek Kadda Wahid0% (1)
- ANALYSE D'UNE Démarche de Cartographie Du RisqueDocument174 pagesANALYSE D'UNE Démarche de Cartographie Du RisqueSouad TouchliftPas encore d'évaluation
- Wayser2009 2Document25 pagesWayser2009 2ALEXANDRE F VOLTAPas encore d'évaluation
- Bac - Les Meilleures Citations Pour Gagner Des Points en Français - SAMABACDocument4 pagesBac - Les Meilleures Citations Pour Gagner Des Points en Français - SAMABACbeavoguipaulbarre047Pas encore d'évaluation
- TD PalettisationDocument12 pagesTD PalettisationZine eddine Hadj mokhnachrPas encore d'évaluation
- Anelise TalbourdeauDocument9 pagesAnelise TalbourdeauDennis AlexanderPas encore d'évaluation
- TABLEAU EDEN ROCK - XLSX 2 VILLADocument20 pagesTABLEAU EDEN ROCK - XLSX 2 VILLARebs RebsPas encore d'évaluation