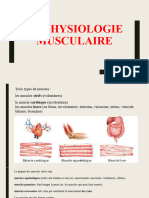
Vous aimerez peut-être aussi
- Muscles: Les Grands Articles d'UniversalisD'EverandMuscles: Les Grands Articles d'UniversalisPas encore d'évaluation
- l3 Neurophysio Chap4Document16 pagesl3 Neurophysio Chap4nobodyPas encore d'évaluation
- Le Muscle SquelletiqueDocument6 pagesLe Muscle SquelletiquefgouzikPas encore d'évaluation
- Cours Myologie PsychomotDocument7 pagesCours Myologie PsychomotRazenAlgPas encore d'évaluation
- P Motricité (C. Quairiaux) - 26.03.21Document5 pagesP Motricité (C. Quairiaux) - 26.03.21nasten16fhPas encore d'évaluation
- Le Fuseau Neuromusculaire-1Document10 pagesLe Fuseau Neuromusculaire-1cristinarotaras2Pas encore d'évaluation
- La MyologieDocument6 pagesLa MyologieBilly Bel Vocale MesidorPas encore d'évaluation
- Le Systeme Musculaire Serie D Exercices Word 1Document11 pagesLe Systeme Musculaire Serie D Exercices Word 1zinebsemlali2009Pas encore d'évaluation
- Controle MedullaireDocument16 pagesControle MedullaireAminou MounirouPas encore d'évaluation
- Physiologie Neuro-MuscuclaireDocument53 pagesPhysiologie Neuro-MuscuclaireMampionona RakotonandrasanaPas encore d'évaluation
- Physiologie de L'appareil Locomoteur-Le Système MusculaireDocument6 pagesPhysiologie de L'appareil Locomoteur-Le Système MusculaireEmy DesreumauxPas encore d'évaluation
- Devoir PhysiologieDocument18 pagesDevoir PhysiologieBetschbel DorcéPas encore d'évaluation
- Syst MusculaireDocument74 pagesSyst Musculaireberthelotsacha85Pas encore d'évaluation
- Le Fuseau Neuromusculaire-1Document10 pagesLe Fuseau Neuromusculaire-1cristinarotaras2Pas encore d'évaluation
- Le Fonctionnement Du Systeme NerveuxDocument11 pagesLe Fonctionnement Du Systeme NerveuxSadi KoukouPas encore d'évaluation
- Chapitre 4 Et Chapitre 5 Muscles Lisse Et Cardiaque.Document14 pagesChapitre 4 Et Chapitre 5 Muscles Lisse Et Cardiaque.habessongPas encore d'évaluation
- Le Muscle Strié Squelettique: Georges CAZORLADocument27 pagesLe Muscle Strié Squelettique: Georges CAZORLAAlphonse AnaniPas encore d'évaluation
- Principes Généraux de La Kinésithérapie 4 (Le Muscle)Document22 pagesPrincipes Généraux de La Kinésithérapie 4 (Le Muscle)8gfm7wjm26Pas encore d'évaluation
- Muscle Et Physiologie MusculaireDocument20 pagesMuscle Et Physiologie MusculaireBoubker PouperPas encore d'évaluation
- Physiologie Musculaire 2020Document47 pagesPhysiologie Musculaire 2020drahlemtrifPas encore d'évaluation
- Circuits NeuronauxDocument23 pagesCircuits NeuronauxPercheryPas encore d'évaluation
- Chapitre 4 Le Systeme MusculaireDocument9 pagesChapitre 4 Le Systeme MusculaireMaxime NowakPas encore d'évaluation
- 2 S7 Cours 2Document44 pages2 S7 Cours 2Chabane OubarechePas encore d'évaluation
- Les MusculesDocument5 pagesLes Musculesnouhailabenallal768Pas encore d'évaluation
- UE 3 Reflexe MedullaireDocument6 pagesUE 3 Reflexe MedullaireCamille DruonPas encore d'évaluation
- Physiologie FCT MusculaireDocument14 pagesPhysiologie FCT MusculaireJon FuagaPas encore d'évaluation
- Les MusclesDocument4 pagesLes Muscleschiha hocinePas encore d'évaluation
- Système Musculaire 2020Document22 pagesSystème Musculaire 2020Sarah OuajnatiPas encore d'évaluation
- La Physiologie MusculaireDocument51 pagesLa Physiologie MusculaireJoronavalona RASAMIMANANAPas encore d'évaluation
- 3 Myologie DEF LightDocument109 pages3 Myologie DEF LightCherbiti Mohammed AminePas encore d'évaluation
- SNC Et SNVDocument3 pagesSNC Et SNVlenoirmccPas encore d'évaluation
- La Contraction MusculaireDocument5 pagesLa Contraction MusculairekharratPas encore d'évaluation
- Sn-Cinesiologie 2021Document3 pagesSn-Cinesiologie 2021jun connexionPas encore d'évaluation
- Chapitre 2 MOUV201Document20 pagesChapitre 2 MOUV201Matéo ROBINPas encore d'évaluation
- L3-CM - Tonus Et Posture 1 - 2023Document38 pagesL3-CM - Tonus Et Posture 1 - 2023DavinaPas encore d'évaluation
- Contraction Musculaire CouleurDocument286 pagesContraction Musculaire Couleurmatis47215100% (1)
- MyologieDocument12 pagesMyologieN'driPas encore d'évaluation
- SN06TEWB0522 Retenir-PartieIDocument10 pagesSN06TEWB0522 Retenir-PartieIremandasarahPas encore d'évaluation
- II Les Réflexes ProprioceptifsDocument31 pagesII Les Réflexes ProprioceptifsHamza Abu Humaid100% (2)
- CM - Composition Et Fonction D'un MuscleDocument3 pagesCM - Composition Et Fonction D'un MuscleBassetPas encore d'évaluation
- SVT - Fiche 14 - RéflexesDocument8 pagesSVT - Fiche 14 - RéflexesClemence BrasseletPas encore d'évaluation
- Système MusculaireDocument16 pagesSystème MusculaireMustapha BriachePas encore d'évaluation
- MuscleDocument15 pagesMuscleMaximePas encore d'évaluation
- 5 La MyologieDocument19 pages5 La MyologieYou NesPas encore d'évaluation
- Annexe Sport Q2Document13 pagesAnnexe Sport Q2descampsPas encore d'évaluation
- Chapitre 5:: Le Système MusculaireDocument5 pagesChapitre 5:: Le Système MusculaireRAYANE YTPas encore d'évaluation
- TSpe T3B1 CoursDocument5 pagesTSpe T3B1 CoursFranckyscocom Gokpan Enfant BeniPas encore d'évaluation
- La Fibre Musculaire TP1Document2 pagesLa Fibre Musculaire TP1salmaessofi35Pas encore d'évaluation
- Contracté - Relâché, Encore Une Fois. - François RosenzweigDocument102 pagesContracté - Relâché, Encore Une Fois. - François RosenzweigOlivia ManceauPas encore d'évaluation
- Cours DAEU 2023 PDFDocument98 pagesCours DAEU 2023 PDFYann PersonnePas encore d'évaluation
- Système Nerveux AutonomeDocument21 pagesSystème Nerveux AutonomeArtichaudPas encore d'évaluation
- Le Syst Me MusculaireDocument27 pagesLe Syst Me MusculaireSarah Dos SantosPas encore d'évaluation
- La Fonction Musculaire Te Le MouvementDocument54 pagesLa Fonction Musculaire Te Le Mouvementmosbahhafida18Pas encore d'évaluation
- 7 - Les Tissus MusculairesDocument6 pages7 - Les Tissus MusculairesHayfa Ben taherPas encore d'évaluation
- Les Tissus MusculairesDocument14 pagesLes Tissus Musculairesariane.vasseur29Pas encore d'évaluation
- Projet EMG (ELECROMYOGRAPHE) FinaliséDocument64 pagesProjet EMG (ELECROMYOGRAPHE) FinaliséAssia Saoudi100% (3)
- BP 2 Boucherie - Le Muscle-CorrectionDocument13 pagesBP 2 Boucherie - Le Muscle-CorrectionThierry Pano Doubleface DucosPas encore d'évaluation
- Poly Physiologie Cellulaire - Contraction Musculaire 2006-2007Document91 pagesPoly Physiologie Cellulaire - Contraction Musculaire 2006-2007roger kinaPas encore d'évaluation
- La MyologieDocument2 pagesLa Myologienouno hohoPas encore d'évaluation
- SVT Le Reflexe Myotatique PDFDocument7 pagesSVT Le Reflexe Myotatique PDFAnonymous TrrvM8Pas encore d'évaluation
- Urgence Chirurgicale de NN 0 L'enfant 2Document32 pagesUrgence Chirurgicale de NN 0 L'enfant 2basmaPas encore d'évaluation
- Tableau Des Sites de Production Des Principales HormonesDocument1 pageTableau Des Sites de Production Des Principales HormonesZineb Dahmani100% (2)
- Cours de Science - La MitoseDocument11 pagesCours de Science - La MitoseArbi SEPas encore d'évaluation
- 3.. Anatomie Et Fonctions de L - App Rep Mâle Élève - DonnéesDocument10 pages3.. Anatomie Et Fonctions de L - App Rep Mâle Élève - DonnéesAbderahmen Haj DahmenPas encore d'évaluation
- Physio RespiratoireDocument31 pagesPhysio RespiratoireOmar Bendriss AlamiPas encore d'évaluation
- Le Systeme Nerveux Central (SNC)Document68 pagesLe Systeme Nerveux Central (SNC)flora.batailleaoustinPas encore d'évaluation
- Les Couleurs en Dermatopathologie: Mise Au PointDocument6 pagesLes Couleurs en Dermatopathologie: Mise Au PointAmina SdPas encore d'évaluation
- Digestion NewDocument245 pagesDigestion Newnaima amina100% (1)
- OssificationDocument25 pagesOssificationBMA-medecinePas encore d'évaluation
- Extrait Du LivreDocument28 pagesExtrait Du LivreYahya RokniPas encore d'évaluation
- Ch. 6: Comportement de Stress, L'adaptabilité de L'organismeDocument2 pagesCh. 6: Comportement de Stress, L'adaptabilité de L'organismeSelda HanimPas encore d'évaluation
- +12. Anémies de L'enfantDocument3 pages+12. Anémies de L'enfantKhawla RiddaPas encore d'évaluation
- 2093 DupasDocument60 pages2093 DupasMohamed Elhadi SaadaPas encore d'évaluation
- 1 - QCM Corrigés de La Lignée Rouge DR ZF PDFDocument23 pages1 - QCM Corrigés de La Lignée Rouge DR ZF PDFMariaPas encore d'évaluation
- Anatomie Et Physiologie de L'edenté TotalDocument9 pagesAnatomie Et Physiologie de L'edenté Totalkedam KedamPas encore d'évaluation
- Série D'exercices N°9 Le Muscle Squelettique 2017 2018 (Mme Harbawi Mbarka)Document6 pagesSérie D'exercices N°9 Le Muscle Squelettique 2017 2018 (Mme Harbawi Mbarka)Hind HindoPas encore d'évaluation
- TissusDocument10 pagesTissusAquarius ManPas encore d'évaluation
- Semiologie de Meridiens CurieuxDocument11 pagesSemiologie de Meridiens CurieuxSoPas encore d'évaluation
- 03 11 Vaisseau Conception REN MAIDocument12 pages03 11 Vaisseau Conception REN MAIKungfudjusuModerneWushuPas encore d'évaluation
- PostersDocument103 pagesPostersHoucine MaghrebiPas encore d'évaluation
- Mémoire de Fin D'études: ThèmeDocument55 pagesMémoire de Fin D'études: ThèmeMordjene GhomariPas encore d'évaluation
- 6-Les Populations Cellulaires LibresDocument50 pages6-Les Populations Cellulaires LibresAy ManPas encore d'évaluation
- Chapitre 4 Anatomie de La FeuilleDocument10 pagesChapitre 4 Anatomie de La FeuilleThe VoidPas encore d'évaluation
- (Ostéopathie) Nouvelle Approche Manipulative. Membre SupérieurDocument519 pages(Ostéopathie) Nouvelle Approche Manipulative. Membre SupérieurRigel Geneste100% (2)
- Groupes Sanguins Modifié S1Document33 pagesGroupes Sanguins Modifié S1Inas HassanatPas encore d'évaluation
- Physiologie Humaine 3eDocument34 pagesPhysiologie Humaine 3eJEAN NDRE100% (1)
- Résumé de La Technique ANAPATH PDFDocument15 pagesRésumé de La Technique ANAPATH PDFAime PrivéPas encore d'évaluation
- Cours Systeme VestibulaireDocument18 pagesCours Systeme VestibulaireMaddy IlPas encore d'évaluation
- 2 - Les Muscles FaciauxDocument19 pages2 - Les Muscles Faciauxfousseni karaPas encore d'évaluation
- 1.anatomie Du Cardio-VasculaireDocument51 pages1.anatomie Du Cardio-VasculaireEmerssone DanPas encore d'évaluation
- Diagnostics différentiels des psychoses endogènes, des troubles de la personnalité et des névrosesD'EverandDiagnostics différentiels des psychoses endogènes, des troubles de la personnalité et des névrosesÉvaluation : 4.5 sur 5 étoiles4.5/5 (2)
- Améliorer votre mémoire: Un Guide pour l'augmentation de la puissance du cerveau, utilisant des techniques et méthodesD'EverandAméliorer votre mémoire: Un Guide pour l'augmentation de la puissance du cerveau, utilisant des techniques et méthodesÉvaluation : 5 sur 5 étoiles5/5 (2)
- Hypnose & affirmations positives pour la richesse, l’abondance et la réussite (21 jours pour un nouveau vous)D'EverandHypnose & affirmations positives pour la richesse, l’abondance et la réussite (21 jours pour un nouveau vous)Évaluation : 5 sur 5 étoiles5/5 (1)
- Guide De Démarrage De L'Apiculture: Le Guide Complet Pour Élever Des AbeillesD'EverandGuide De Démarrage De L'Apiculture: Le Guide Complet Pour Élever Des AbeillesPas encore d'évaluation
- Neuropsychologie: Les bases théoriques et pratiques du domaine d'étude (psychologie pour tous)D'EverandNeuropsychologie: Les bases théoriques et pratiques du domaine d'étude (psychologie pour tous)Pas encore d'évaluation
- Le Pouvoir De L'Autodiscipline: L’art de tout accomplir sans limiteD'EverandLe Pouvoir De L'Autodiscipline: L’art de tout accomplir sans limiteÉvaluation : 4 sur 5 étoiles4/5 (16)
- La vie des abeilles: Prix Nobel de littératureD'EverandLa vie des abeilles: Prix Nobel de littératureÉvaluation : 4 sur 5 étoiles4/5 (41)
- Introduction à la psychologie des émotions: De Darwin aux neurosciences: découvrir les émotions et leur mode de fonctionnementD'EverandIntroduction à la psychologie des émotions: De Darwin aux neurosciences: découvrir les émotions et leur mode de fonctionnementPas encore d'évaluation
- Harmonisation Energétique des Lieux: Habitat et haut-lieux sacrés 2020D'EverandHarmonisation Energétique des Lieux: Habitat et haut-lieux sacrés 2020Évaluation : 2.5 sur 5 étoiles2.5/5 (3)
- Histoire de la psychologie scientifique: De la naissance de la psychologie à la neuropsychologie et aux champs d'application les plus actuelsD'EverandHistoire de la psychologie scientifique: De la naissance de la psychologie à la neuropsychologie et aux champs d'application les plus actuelsPas encore d'évaluation
- Géologie de l'Amérique: Les Grands Articles d'UniversalisD'EverandGéologie de l'Amérique: Les Grands Articles d'UniversalisPas encore d'évaluation
- La psychologie de l'anxiété: La connaître pour comprendre ses mécanismes de fonctionnementD'EverandLa psychologie de l'anxiété: La connaître pour comprendre ses mécanismes de fonctionnementPas encore d'évaluation
- Cahier de jeux de stimulation cognitive: Sujets Alzheimer, désorientés, démences, amnésiesD'EverandCahier de jeux de stimulation cognitive: Sujets Alzheimer, désorientés, démences, amnésiesPas encore d'évaluation
- La Conscience Et L'Univers Existent Sans Commencement Ni FinD'EverandLa Conscience Et L'Univers Existent Sans Commencement Ni FinPas encore d'évaluation
- Transition énergétique ET Changement climatique: Développements et Perspectives d'AvenirD'EverandTransition énergétique ET Changement climatique: Développements et Perspectives d'AvenirPas encore d'évaluation
- Le CODE DE DIEU: Le secret de notre passé, la promesse de notre avenirD'EverandLe CODE DE DIEU: Le secret de notre passé, la promesse de notre avenirÉvaluation : 4 sur 5 étoiles4/5 (23)
- Le TDAH chez l’adulte : Comment reconnaître & faire face au TDAH chez l’adulte en 30 étapes faciles.D'EverandLe TDAH chez l’adulte : Comment reconnaître & faire face au TDAH chez l’adulte en 30 étapes faciles.Pas encore d'évaluation
- Hypersensible Et alors ?: Un tempérament expliquéD'EverandHypersensible Et alors ?: Un tempérament expliquéÉvaluation : 5 sur 5 étoiles5/5 (2)
- Autisme - La boîte à outils: Stratégies et techniques pour accompagner un enfant autisteD'EverandAutisme - La boîte à outils: Stratégies et techniques pour accompagner un enfant autistePas encore d'évaluation
- Qu'arrive t'il a la terre arable ?: Le problème de l'érosion du sol.D'EverandQu'arrive t'il a la terre arable ?: Le problème de l'érosion du sol.Évaluation : 5 sur 5 étoiles5/5 (1)