Vous aimerez peut-être aussi
- Influx Nerveux 2Document22 pagesInflux Nerveux 2Samia Belabbas100% (1)
- Cours ImagerieDocument19 pagesCours ImagerieKarimaKarimaPas encore d'évaluation
- Mécanique de Fluide - by YoupaaaDocument9 pagesMécanique de Fluide - by YoupaaaAmel Marisol SahagùnPas encore d'évaluation
- Le Condensateur Bac Sc.M.T.inf.-ConvertiDocument6 pagesLe Condensateur Bac Sc.M.T.inf.-ConvertiBenmohamed ibtihelPas encore d'évaluation
- Correction TD 7 - 4P050Document10 pagesCorrection TD 7 - 4P050badid SusskindPas encore d'évaluation
- 0 TD Fso Liaison Chimique 15 16Document20 pages0 TD Fso Liaison Chimique 15 16Ursule YaméogoPas encore d'évaluation
- Cryptographie QuantiqueDocument46 pagesCryptographie Quantiquebenaouda310% (1)
- CEMS Physique 2015 WahabdiopDocument7 pagesCEMS Physique 2015 WahabdiopAida MbodjiPas encore d'évaluation
- OLYMPIADE - Niveau PremièreDocument14 pagesOLYMPIADE - Niveau PremièrealfredPas encore d'évaluation
- Maths 2C - L12 - HomothétieDocument14 pagesMaths 2C - L12 - Homothétieben danPas encore d'évaluation
- RC1 1Document5 pagesRC1 1mehdi benmassoudPas encore d'évaluation
- La Mécanique de Newton Étude de Cas PDFDocument10 pagesLa Mécanique de Newton Étude de Cas PDFMarøøen Bouabid100% (1)
- Annal MathDocument53 pagesAnnal MathLotfi Touil100% (1)
- TD Magnetos - L2-PC-EEM-EEAI-2022-2023 Partie 2 - 230322 - 174738Document5 pagesTD Magnetos - L2-PC-EEM-EEAI-2022-2023 Partie 2 - 230322 - 174738Axel AssamoaPas encore d'évaluation
- Série 1Document2 pagesSérie 1Japhet DOSSOUPas encore d'évaluation
- A2 Niveaux D Energie AtomesDocument11 pagesA2 Niveaux D Energie AtomesfaslaPas encore d'évaluation
- Composet 11Document2 pagesComposet 11Oumar TraoréPas encore d'évaluation
- TD N04 Theorie Des Champs-1Document2 pagesTD N04 Theorie Des Champs-1M'pounguiPas encore d'évaluation
- Induction Lorentz ExosDocument4 pagesInduction Lorentz ExosLilia BennaceurPas encore d'évaluation
- Chap 3Document24 pagesChap 3Ayoub BPas encore d'évaluation
- Algorithmique Et Programmation - Grintta - Bot PDFDocument56 pagesAlgorithmique Et Programmation - Grintta - Bot PDFtouati souadPas encore d'évaluation
- Matrices Et Déterminants - Généralités Sur Les MatricesDocument3 pagesMatrices Et Déterminants - Généralités Sur Les MatricesHamza PrintoOsPas encore d'évaluation
- Lois de Composition Interne Corrige Serie D Exercices 1Document10 pagesLois de Composition Interne Corrige Serie D Exercices 1Taha El KouitPas encore d'évaluation
- Tseba 2021Document3 pagesTseba 2021Dado DjiguibaPas encore d'évaluation
- RDP - Notions de Sciences Physiques Et NaturellesDocument578 pagesRDP - Notions de Sciences Physiques Et Naturellesbelgam20% (1)
- Mouvement Dans Un Champ Uniforme - Exercices - Devoirs: Exercice 1Document9 pagesMouvement Dans Un Champ Uniforme - Exercices - Devoirs: Exercice 1Amine AlaoUii Alaoui100% (1)
- DevoirDocument2 pagesDevoirOusseynou DiagnePas encore d'évaluation
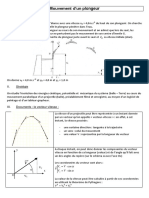
- Mouvement D Un PlongeurDocument5 pagesMouvement D Un PlongeurmounirPas encore d'évaluation
- Physique TleC TD College Henri Dumont 2019 2020Document4 pagesPhysique TleC TD College Henri Dumont 2019 2020HENRIETTEPas encore d'évaluation
- ANNALESBAC2011Document42 pagesANNALESBAC2011charlesmartel55100% (1)
- Cours 2 Les Systèmes Mécaniques OscillantsDocument121 pagesCours 2 Les Systèmes Mécaniques OscillantsMorgan merdaPas encore d'évaluation
- TD PolynomeDocument1 pageTD Polynomeeric100% (1)
- SMPC s2 CHP 2Document31 pagesSMPC s2 CHP 2Abdelhakim BailalPas encore d'évaluation
- TD Magnetos - L2-PC-EEM-EEAI-2022-2023 Partie 1 (1) - 230329 - 163846Document3 pagesTD Magnetos - L2-PC-EEM-EEAI-2022-2023 Partie 1 (1) - 230329 - 163846Axel AssamoaPas encore d'évaluation
- P6-5-Rayonnement D Un Dipole Oscillant PDFDocument11 pagesP6-5-Rayonnement D Un Dipole Oscillant PDFHamma MeskiniPas encore d'évaluation
- Revisions Algebre LineaireDocument53 pagesRevisions Algebre Lineairebestgaming player100% (1)
- 2-Ondes Électromagnétiques Dans Un Conducteur Et Dans Un PlasmaDocument9 pages2-Ondes Électromagnétiques Dans Un Conducteur Et Dans Un PlasmaNoureddine BoxePas encore d'évaluation
- Thermo Stat PBDocument61 pagesThermo Stat PBabdelkanPas encore d'évaluation
- Sujet Bac TSE Physique 2023Document3 pagesSujet Bac TSE Physique 2023Toumany FofanaPas encore d'évaluation
- Ds 1 MP 2022Document3 pagesDs 1 MP 2022mehdi benmassoudPas encore d'évaluation
- Sujet de Révision N°1 (Corrigé) - Maths - Bac Sciences (2009-2010) MR Abdelbasset LaataouiDocument7 pagesSujet de Révision N°1 (Corrigé) - Maths - Bac Sciences (2009-2010) MR Abdelbasset LaataouiBessem ChalloufPas encore d'évaluation
- Polynomes Cours ENSADocument11 pagesPolynomes Cours ENSAyassine laarbiPas encore d'évaluation
- CH16 Temps Et Relativité Restreinte PDFDocument4 pagesCH16 Temps Et Relativité Restreinte PDFanon_819903036Pas encore d'évaluation
- TSE Maths BAC2014Document3 pagesTSE Maths BAC2014Adama MaigaPas encore d'évaluation
- 03 Electrocinetique Lois Generales ARQS Exercices PDFDocument3 pages03 Electrocinetique Lois Generales ARQS Exercices PDFlasso cissePas encore d'évaluation
- PC1ere CD - P6,5 Champ ElestrostatiqueDocument9 pagesPC1ere CD - P6,5 Champ ElestrostatiqueAdopo100% (1)
- Etude D'un Dipole (R, L)Document6 pagesEtude D'un Dipole (R, L)Mohamed Soibaha ChaambanePas encore d'évaluation
- Outils Mathématique Pour PhysiqueDocument22 pagesOutils Mathématique Pour Physiqueabdelouahab bouchama100% (1)
- 10 Cours Relativite Du TempsDocument7 pages10 Cours Relativite Du TempsCéline LuttiPas encore d'évaluation
- Apc Cours de Physiques TD Et TC.Document73 pagesApc Cours de Physiques TD Et TC.Magatte DiopPas encore d'évaluation
- TD N3 DynamiqueDocument2 pagesTD N3 DynamiqueSara Akli100% (1)
- Fax TDDocument40 pagesFax TDjpwhwdpbytPas encore d'évaluation
- 03 Exercices P4 Projection ForcesDocument1 page03 Exercices P4 Projection ForcesSiham YahansalPas encore d'évaluation
- Dyn en ExoDocument11 pagesDyn en ExodarkparadizPas encore d'évaluation
- CorrectionDocument8 pagesCorrectiondriss ouardiPas encore d'évaluation
- Enseignement AstronomieDocument184 pagesEnseignement AstronomiebougetPas encore d'évaluation
- Table HistoireDocument3 pagesTable HistoirebougetPas encore d'évaluation
- Riverin Celine 2014 TheseDocument235 pagesRiverin Celine 2014 ThesebougetPas encore d'évaluation
- Glasgow Art NouveauDocument3 pagesGlasgow Art NouveaubougetPas encore d'évaluation
- ROGER LAMOULINE-Le Telescope Disaac Newton Et Moi - (Atramenta - Net)Document16 pagesROGER LAMOULINE-Le Telescope Disaac Newton Et Moi - (Atramenta - Net)bougetPas encore d'évaluation
- BA2b Cours05 KuhnDocument34 pagesBA2b Cours05 KuhnbougetPas encore d'évaluation
- Rea 0035-2004 1988 Num 90 1 4326Document7 pagesRea 0035-2004 1988 Num 90 1 4326bougetPas encore d'évaluation
- Une Merveille Oubliee LastrolabeDocument28 pagesUne Merveille Oubliee LastrolabebougetPas encore d'évaluation
- La Lune Au Secours Des Marins - La Déconvenue D'alexis ClairautDocument7 pagesLa Lune Au Secours Des Marins - La Déconvenue D'alexis ClairautbougetPas encore d'évaluation
- 8.python Date HeureDocument7 pages8.python Date HeureNourchene BensmayaPas encore d'évaluation
- Guide PoissonDocument2 pagesGuide PoissonphypaPas encore d'évaluation
- Latin TechniqueDocument16 pagesLatin TechniquebougetPas encore d'évaluation
- Cours TD Phys 601 PCDocument56 pagesCours TD Phys 601 PCbougetPas encore d'évaluation
- Dispute Sur La Parallaxe de La LuneDocument4 pagesDispute Sur La Parallaxe de La LunebougetPas encore d'évaluation
- Villa Alessandro p05 FlattenDocument39 pagesVilla Alessandro p05 FlattenFranck BitaPas encore d'évaluation
- Vuillez Jean Philippe p01Document50 pagesVuillez Jean Philippe p01bougetPas encore d'évaluation
- Villa Alessandro p1Document30 pagesVilla Alessandro p1bougetPas encore d'évaluation
- Stellarium Scripts enDocument5 pagesStellarium Scripts enbougetPas encore d'évaluation
- Villa Alessandro p4Document59 pagesVilla Alessandro p4bougetPas encore d'évaluation
- Villa Alessandro p02Document58 pagesVilla Alessandro p02bougetPas encore d'évaluation
- Pebay Peyroula Eva p00Document8 pagesPebay Peyroula Eva p00bougetPas encore d'évaluation
- Pebay Peyroula Eva p02 PDFDocument32 pagesPebay Peyroula Eva p02 PDFAissamErraPas encore d'évaluation
- Pebay Peyroula Eva p03Document32 pagesPebay Peyroula Eva p03مصطفاوي محمدPas encore d'évaluation
- Pebay Peyroula Eva p04Document28 pagesPebay Peyroula Eva p04bougetPas encore d'évaluation
- Pebay Peyroula Eva p01Document21 pagesPebay Peyroula Eva p01bougetPas encore d'évaluation
- Pebay Peyroula Eva p08Document36 pagesPebay Peyroula Eva p08Yahya AidaraPas encore d'évaluation
- Pebay Peyroula Eva p05Document33 pagesPebay Peyroula Eva p05malikPas encore d'évaluation
- Pebay Peyroula Eva p07Document25 pagesPebay Peyroula Eva p07bougetPas encore d'évaluation
- Pebay Peyroula Eva p06Document28 pagesPebay Peyroula Eva p06Mohammed ChamiPas encore d'évaluation
- DM PhysiqueDocument5 pagesDM Physiqueالقي السمع وٱفتح قلبكPas encore d'évaluation
- Additifs AlimentairesDocument56 pagesAdditifs AlimentairesI. Ez100% (3)
- OIE Principes Generaux DesinfectionDocument20 pagesOIE Principes Generaux DesinfectionMustapha 471989Pas encore d'évaluation
- PolutionDocument5 pagesPolutionsid mebarekPas encore d'évaluation
- Cours GlucidesDocument66 pagesCours GlucidesAli RymePas encore d'évaluation
- U31 - 2009 - PhyschimDocument8 pagesU31 - 2009 - PhyschimHamza OuniPas encore d'évaluation
- Exposé Du Soudage Au LaserDocument37 pagesExposé Du Soudage Au Laserassoua kloliè konanPas encore d'évaluation
- Pétrole Et GazDocument8 pagesPétrole Et GazAMADOU DOUMBIAPas encore d'évaluation
- SousouDocument3 pagesSousousoumeya soltaniPas encore d'évaluation
- 02 Nomenclature2004Document95 pages02 Nomenclature2004Michel Tolima kitokoPas encore d'évaluation
- Pollution Généres Par Les Déchets Solides Et Les Techniques D'atténuation Et Des Réduction Des Nuissances PDFDocument60 pagesPollution Généres Par Les Déchets Solides Et Les Techniques D'atténuation Et Des Réduction Des Nuissances PDFOumaima Ncib100% (1)
- Fuite de Chlorure de Vinyle Et de PVC Dans Une Usine ChimiqueDocument8 pagesFuite de Chlorure de Vinyle Et de PVC Dans Une Usine Chimiqueagha hutamaPas encore d'évaluation
- Exercice Chimie BacDocument2 pagesExercice Chimie BacAli HaniniPas encore d'évaluation
- Cours - Fabrication Et Transformation Des Polymères - pp47-81Document35 pagesCours - Fabrication Et Transformation Des Polymères - pp47-81atefdinPas encore d'évaluation
- Programme 1. Licence Analytique Programme Détaillé Par Matière Des Semestres S5 Et S6Document44 pagesProgramme 1. Licence Analytique Programme Détaillé Par Matière Des Semestres S5 Et S6Abdou HkPas encore d'évaluation
- Cours BétonDocument5 pagesCours BétonOuezna BahloulPas encore d'évaluation
- EMI-S2-1GC-ELBRAHMI-S2T3-Géotechnique 1Document54 pagesEMI-S2-1GC-ELBRAHMI-S2T3-Géotechnique 1Ghi Tta50% (2)
- Flopam An 905 VHM (MC) - Can - CFDocument9 pagesFlopam An 905 VHM (MC) - Can - CFAlain gPas encore d'évaluation
- Chap Extraction Ration Esp ChimiquesتDocument6 pagesChap Extraction Ration Esp ChimiquesتMouhibi AbdellahPas encore d'évaluation
- Rapport PFEDocument106 pagesRapport PFETamis HamzaPas encore d'évaluation
- 3 Bouabdallah2-Sngc07Document9 pages3 Bouabdallah2-Sngc07Mohamed HouhouPas encore d'évaluation
- Chimie Pharmaceutique IDocument57 pagesChimie Pharmaceutique ITadjadine AbdoulayePas encore d'évaluation
- TD 2 Et 3Document9 pagesTD 2 Et 3ahmedPas encore d'évaluation
- 03 001 GlametDocument4 pages03 001 GlametNora MazariPas encore d'évaluation
- TP Toxicologie Pharmacie 3 ADocument11 pagesTP Toxicologie Pharmacie 3 Afatima ezzahrae zananiPas encore d'évaluation
- MMP G3 2020Document67 pagesMMP G3 2020CharlesPas encore d'évaluation
- Série Solutions Acides Et Basiques Prof - Elhabib (WWW - Pc1.ma)Document2 pagesSérie Solutions Acides Et Basiques Prof - Elhabib (WWW - Pc1.ma)Agnaou MohamedPas encore d'évaluation
- Fiches Produits FR 2Document104 pagesFiches Produits FR 2pauline dennemontPas encore d'évaluation
- CH 1Document15 pagesCH 1Tech KingPas encore d'évaluation
- C. Biochimie III. Lipides (III.4. Métabolisme Du Cholestérol)Document26 pagesC. Biochimie III. Lipides (III.4. Métabolisme Du Cholestérol)Rania HamdiPas encore d'évaluation