Vous aimerez peut-être aussi
- CHAPITRE 4 Immunité InnéeDocument13 pagesCHAPITRE 4 Immunité InnéeJuan Sebastian MoraPas encore d'évaluation
- Capture d’écran . 2024-03-20 à 17.34.52Document16 pagesCapture d’écran . 2024-03-20 à 17.34.52pce.chloePas encore d'évaluation
- ImmunoDocument29 pagesImmunoIkram DahmaniPas encore d'évaluation
- La Réponse Immunitaire Innée Et Adaptative ModifDocument8 pagesLa Réponse Immunitaire Innée Et Adaptative ModifAni TrghwiPas encore d'évaluation
- Chapitre 1 - Introduction À L - Immunologie - ElumensDocument9 pagesChapitre 1 - Introduction À L - Immunologie - ElumenstructresdouteuxPas encore d'évaluation
- SVT Fiche RevisionsDocument1 pageSVT Fiche RevisionsClothilde LaforêtPas encore d'évaluation
- Infectiologie - Roneo 18.01.11Document9 pagesInfectiologie - Roneo 18.01.11nicolas43Pas encore d'évaluation
- Cours Histologie 2 Partie 2Document66 pagesCours Histologie 2 Partie 2Salomon JosephPas encore d'évaluation
- Le Système ImmunitaireDocument18 pagesLe Système ImmunitaireccPas encore d'évaluation
- ImmunoDocument5 pagesImmunoAmadou CISSEPas encore d'évaluation
- Immunologie - Cahier PRDocument64 pagesImmunologie - Cahier PRJulie LEMOINEPas encore d'évaluation
- Cours 1 d'Immuno2LSP23Document27 pagesCours 1 d'Immuno2LSP23abdou2004trPas encore d'évaluation
- CytokinesDocument6 pagesCytokinesfsxnsstoaehkbhlzygPas encore d'évaluation
- Immuno 1Document4 pagesImmuno 1Rkia EddabraPas encore d'évaluation
- Le Systeme ImmunitaireDocument32 pagesLe Systeme ImmunitaireIlyes MraidiaPas encore d'évaluation
- Introduction ImmunologieDocument29 pagesIntroduction Immunologieagag.salahPas encore d'évaluation
- Recuerdo Fisiológico Del Sistema Inmunológico - FR CorregidoDocument9 pagesRecuerdo Fisiológico Del Sistema Inmunológico - FR CorregidoIbrahima.senghor KalissaPas encore d'évaluation
- Le Systeme Immunitaire Cours FRDocument3 pagesLe Systeme Immunitaire Cours FR1BAC SVT1Pas encore d'évaluation
- L'Immunologie - SVT 1èrea Tome2Document7 pagesL'Immunologie - SVT 1èrea Tome2Guy TangaPas encore d'évaluation
- Cours D'immunologie Médicale PDFDocument17 pagesCours D'immunologie Médicale PDFMafankoloPas encore d'évaluation
- Mécanisme de Défense SpécifiqueDocument6 pagesMécanisme de Défense SpécifiqueJoseph OnanaPas encore d'évaluation
- cours immunologie S5 (2)Document113 pagescours immunologie S5 (2)hamzaabzou154Pas encore d'évaluation
- Essentiel11 Immunite AdaptativeDocument2 pagesEssentiel11 Immunite Adaptativejad mamouPas encore d'évaluation
- MÉCANISME DE LA RÉPONSE IMMUNITAIRE – Senrevision.comDocument12 pagesMÉCANISME DE LA RÉPONSE IMMUNITAIRE – Senrevision.comKurtisgangue18Pas encore d'évaluation
- RappelDocument11 pagesRappelNadjib A. NedjarPas encore d'évaluation
- Cours ImmunoDocument29 pagesCours ImmunoChristian NgalaPas encore d'évaluation
- Les Réponses Immunitaires DR MILOUDIDocument13 pagesLes Réponses Immunitaires DR MILOUDIFarah B. BtoushPas encore d'évaluation
- BIOS 308 IMMUNOLOGIE GENERALE ET FONCTIONELLE (1)Document159 pagesBIOS 308 IMMUNOLOGIE GENERALE ET FONCTIONELLE (1)Mahamat Mahamat AboubakarPas encore d'évaluation
- L'Immunité SpécifiqueDocument12 pagesL'Immunité Spécifiqueraslenbouallegue12345Pas encore d'évaluation
- Immunite CoursDocument14 pagesImmunite CoursJulien GhandourPas encore d'évaluation
- المستندDocument4 pagesالمستندlakhfksnufPas encore d'évaluation
- 1 Er Cours Immuno Appl BS 202 PDFDocument10 pages1 Er Cours Immuno Appl BS 202 PDFHasna BentPas encore d'évaluation
- Thème 3 Chapitre 3Document27 pagesThème 3 Chapitre 3TIMCCPas encore d'évaluation
- COURSDocument3 pagesCOURSzeksmdlw lsqd;flc;wùPas encore d'évaluation
- Organes lymphoidesAMROUNDocument46 pagesOrganes lymphoidesAMROUNAya BoucharebPas encore d'évaluation
- Bilan Chapitre 3 L ' Immunite AdaptativeDocument6 pagesBilan Chapitre 3 L ' Immunite Adaptativeahmedrayanelali23Pas encore d'évaluation
- Systeme ImmunitaireDocument31 pagesSysteme Immunitairehadji0% (1)
- 02 - La Reponse Inflammatoire - Interactions Leucocytes EndothéliumDocument6 pages02 - La Reponse Inflammatoire - Interactions Leucocytes EndothéliumMohamed-Amine SADKIPas encore d'évaluation
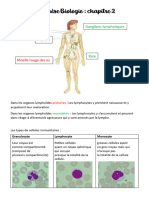
- Synthèse Biologie chp2Document6 pagesSynthèse Biologie chp2clara.plv22Pas encore d'évaluation
- QUELQUES NOTIONS DU COURS DDocument2 pagesQUELQUES NOTIONS DU COURS Dazaraadama755Pas encore d'évaluation
- Chapitre II Organes Et Cellules de L Immunit PDF PDFDocument22 pagesChapitre II Organes Et Cellules de L Immunit PDF PDFسارة عبيديPas encore d'évaluation
- Fiche IntroDocument3 pagesFiche IntrorequiemforathrowawayPas encore d'évaluation
- Physiopathologie de LDocument10 pagesPhysiopathologie de LikramPas encore d'évaluation
- Ronéos HématologieDocument123 pagesRonéos HématologieMohamed-Amine SADKIPas encore d'évaluation
- Immunologie 1Document52 pagesImmunologie 1Mustafa KouzritePas encore d'évaluation
- Immuno3an-Reponses Immunitaires2018benachourDocument9 pagesImmuno3an-Reponses Immunitaires2018benachourAmina MnPas encore d'évaluation
- Introduction Générale À LimmunologieDocument67 pagesIntroduction Générale À LimmunologieNabil holmes100% (1)
- Anapath3an-Processus Inflammatoire2022benahseneDocument10 pagesAnapath3an-Processus Inflammatoire2022benahseneouss said100% (1)
- 3eme TH3 Immuno E2 Réponse GlobaleDocument2 pages3eme TH3 Immuno E2 Réponse Globalet5fr62fgshPas encore d'évaluation
- 1 - Introduction À L'immunologieDocument4 pages1 - Introduction À L'immunologieOsama O-bPas encore d'évaluation
- Système ImmunitaireDocument34 pagesSystème ImmunitaireAmeen MaoPas encore d'évaluation
- Systeme ImmunologiqueDocument8 pagesSysteme ImmunologiqueAhmed MarwanPas encore d'évaluation
- Questions Système Immunitaire CorrigéDocument3 pagesQuestions Système Immunitaire CorrigéWou SOUAREPas encore d'évaluation
- 02-immunitée-innée-24-Document11 pages02-immunitée-innée-24-sarahchawech68Pas encore d'évaluation
- Chapitre III Cours Immunologie SVT3Document26 pagesChapitre III Cours Immunologie SVT3سالم شعبانPas encore d'évaluation
- Chapitre 1. Introduction Au Système ImmunitaireDocument69 pagesChapitre 1. Introduction Au Système Immunitairefallone zannouPas encore d'évaluation
- Immuno 2Document28 pagesImmuno 2Ameen MaoPas encore d'évaluation
- Les cellules immunitairesDocument23 pagesLes cellules immunitairesMoundher HafsiPas encore d'évaluation
- ImmunologieDocument24 pagesImmunologieayoub berrePas encore d'évaluation
- Amélie NothombDocument9 pagesAmélie NothombGi EmmePas encore d'évaluation
- Rapport Projet - Aménagement Du TerritoireDocument22 pagesRapport Projet - Aménagement Du TerritoireGRCStudent CommitteePas encore d'évaluation
- Exposé RhizomanieDocument18 pagesExposé RhizomanieSimo DadsiPas encore d'évaluation
- Vivacité N°3 - Septembre-Octobre 2009 - Les MureauxDocument20 pagesVivacité N°3 - Septembre-Octobre 2009 - Les MureauxMairie des MureauxPas encore d'évaluation
- Durabilité Des Aliments Pour Le Poisson en AquacultureDocument17 pagesDurabilité Des Aliments Pour Le Poisson en AquacultureSabour IssaPas encore d'évaluation
- EXERCICES RESOLUS Concentration Solutions ÉlectrolytiquesDocument8 pagesEXERCICES RESOLUS Concentration Solutions ÉlectrolytiquesKenza TouizaPas encore d'évaluation
- Diagnostics IncroyablesDocument414 pagesDiagnostics IncroyablesHICHAM HACHLAFPas encore d'évaluation
- Bruno Clavier - Les Fantômes de L'analysteDocument309 pagesBruno Clavier - Les Fantômes de L'analysteJuju Studio SarahPas encore d'évaluation
- 5563Document17 pages5563Ali GouriPas encore d'évaluation
- Détecteurs Et Émetteurs Optoélectroniques À Semi-Conducteurs (Enregistrement Automatique)Document81 pagesDétecteurs Et Émetteurs Optoélectroniques À Semi-Conducteurs (Enregistrement Automatique)Ahlam BOUANIPas encore d'évaluation
- Jeu Et Réalité Lespace Potentiel (Donald Winnicott) (Z-Library)Document222 pagesJeu Et Réalité Lespace Potentiel (Donald Winnicott) (Z-Library)inouPas encore d'évaluation
- th0238066 TheseDocument231 pagesth0238066 ThesePFEPas encore d'évaluation
- Dosages Acide ConvertiDocument6 pagesDosages Acide ConvertiAbir SoujaaPas encore d'évaluation
- Lexium SD3 - LU9GC3Document2 pagesLexium SD3 - LU9GC3Mohamed Amine LABIDIPas encore d'évaluation
- Formation Réticulaire (Thèse)Document117 pagesFormation Réticulaire (Thèse)Franck DernoncourtPas encore d'évaluation
- 2015 ELALAOUI Diffusion PDFDocument215 pages2015 ELALAOUI Diffusion PDFRou MaissaPas encore d'évaluation
- HPV PDFDocument3 pagesHPV PDFAnonymous VbEvDIOUaPas encore d'évaluation
- 1633 20150316 PDFDocument20 pages1633 20150316 PDFelmoudjahid_dzPas encore d'évaluation
- Physiologie: MédicaleDocument88 pagesPhysiologie: MédicaleSganarelle St hubertPas encore d'évaluation
- Regime TNDocument21 pagesRegime TNĐăng HuyPas encore d'évaluation
- 20-00511-Centrale Danone-RF2019 22janvier 1 CompressedDocument62 pages20-00511-Centrale Danone-RF2019 22janvier 1 CompressedjadPas encore d'évaluation
- t4 Fill 23eDocument2 pagest4 Fill 23eAdemuyiwa OlaniyiPas encore d'évaluation
- Fait Divers 3PDocument14 pagesFait Divers 3PFarid FaridPas encore d'évaluation
- Presentation Generale RBPP Sante Mineurs Jeunes MajeursDocument12 pagesPresentation Generale RBPP Sante Mineurs Jeunes Majeursfranck hermann tapePas encore d'évaluation
- Stanke DESC PharmacologieDocument40 pagesStanke DESC PharmacologieKawther BoudifaPas encore d'évaluation
- Chap10 Orthomyxoviridae PDFDocument30 pagesChap10 Orthomyxoviridae PDFDiopPas encore d'évaluation
- Rapport de StageDocument7 pagesRapport de StageZïnbē MėäãmërPas encore d'évaluation
- Lenovo-Pc20200624145014 PDFDocument3 pagesLenovo-Pc20200624145014 PDFMOUMOUPas encore d'évaluation