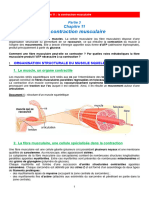
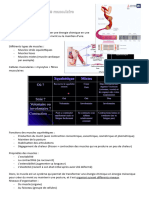
Vous aimerez peut-être aussi
- La Contraction MusculaireDocument6 pagesLa Contraction MusculaireLim BunkimPas encore d'évaluation
- La Contraction MusculaireDocument11 pagesLa Contraction MusculaireJihadi AbbouPas encore d'évaluation
- 4.3.2.mécanismes de La Contractio1Document4 pages4.3.2.mécanismes de La Contractio1Ayoub LakhdariPas encore d'évaluation
- Electro - Syst Nerveux - CHAP IV B Transmission NeuromusculaireDocument68 pagesElectro - Syst Nerveux - CHAP IV B Transmission NeuromusculaireAliou LabaykaPas encore d'évaluation
- 1-L'Excitation Musculaire Libère deDocument1 page1-L'Excitation Musculaire Libère deNizar SaadPas encore d'évaluation
- NEUROPHYSIOLOGIEDocument47 pagesNEUROPHYSIOLOGIEPrince AlegaPas encore d'évaluation
- Physiologie Du Muscle Et de La Contraction MusculaireDocument5 pagesPhysiologie Du Muscle Et de La Contraction MusculaireImane BenderazPas encore d'évaluation
- Diapo Physio Hennock-2Document31 pagesDiapo Physio Hennock-2mossila BradleyPas encore d'évaluation
- Correction de La Série N°9 - SVT - Muscle Squelettique - Bac Sciences Exp (2018-2019) Mme Mbarka HarbawiDocument10 pagesCorrection de La Série N°9 - SVT - Muscle Squelettique - Bac Sciences Exp (2018-2019) Mme Mbarka HarbawiSassi Lassaad100% (1)
- Activité éléctrique Du Coeur (Dã©but)Document13 pagesActivité éléctrique Du Coeur (Dã©but)cc44vnzh4kPas encore d'évaluation
- Physiologie Muscles Striés SquelettiquesDocument16 pagesPhysiologie Muscles Striés SquelettiquessafemindPas encore d'évaluation
- Muscles Cours2Document3 pagesMuscles Cours2fardadi rachidPas encore d'évaluation
- Les Tissus MusculairesDocument11 pagesLes Tissus MusculairesZoulkiffiPas encore d'évaluation
- Khaiter Devoir Numero 1Document5 pagesKhaiter Devoir Numero 1Y2kMagiikPas encore d'évaluation
- ch10 Solutionnaire 1569538426Document12 pagesch10 Solutionnaire 1569538426Wilfried KouaoPas encore d'évaluation
- Physiologie Du Muscle Strie SquelettiqueDocument39 pagesPhysiologie Du Muscle Strie SquelettiqueRaouf ChikhiPas encore d'évaluation
- Physio1an31-7jonction NeuromusculaireDocument15 pagesPhysio1an31-7jonction Neuromusculaireelidrissimaryam733Pas encore d'évaluation
- Le Muscle SquelettiqueDocument4 pagesLe Muscle SquelettiqueGabriela TituPas encore d'évaluation
- Physiologie LA JONCTION NEUROMUSCULAIREDocument9 pagesPhysiologie LA JONCTION NEUROMUSCULAIREtoni.slmPas encore d'évaluation
- 2-Physiologie MusculaireDocument44 pages2-Physiologie MusculaireIslęm OuaribPas encore d'évaluation
- CM Muscle FRB 6 DiaDocument13 pagesCM Muscle FRB 6 DiaWilly Le DruillennecPas encore d'évaluation
- Les Muscles LissesDocument5 pagesLes Muscles LissesHassanElhassanPas encore d'évaluation
- Cours 1 CytosqueletteDocument12 pagesCours 1 CytosquelettenadeauPas encore d'évaluation
- Hand Physiologie MuscleDocument9 pagesHand Physiologie MuscleRachid BenPas encore d'évaluation
- Activité MsculaireDocument7 pagesActivité MsculaireameniPas encore d'évaluation
- 4.contraction MusculaireDocument40 pages4.contraction MusculaireKima MadPas encore d'évaluation
- Chapitre4.1 Biochimie Cellulaire Et FonctionnelleDocument7 pagesChapitre4.1 Biochimie Cellulaire Et Fonctionnelledraboissouf174Pas encore d'évaluation
- Contraction MusculaireDocument1 pageContraction MusculaireZoha AboudarghamPas encore d'évaluation
- VasodilatadoresDocument16 pagesVasodilatadoresGloria PotellaPas encore d'évaluation
- La Contraction Musculaire PDFDocument9 pagesLa Contraction Musculaire PDFFaculté De Médecine Béchar50% (2)
- 08-Physiologie de La Jonction NeuromusculaireDocument26 pages08-Physiologie de La Jonction NeuromusculaireBook HunterPas encore d'évaluation
- 03 - 2 - Proprietes Mecaniques Du Muscle - ContractionDocument30 pages03 - 2 - Proprietes Mecaniques Du Muscle - ContractionGloria KitonPas encore d'évaluation
- Bilan Du Chapitre 11 - La Contraction MusculaireDocument12 pagesBilan Du Chapitre 11 - La Contraction MusculaireYousra INDIAPas encore d'évaluation
- Physiologie Du Mucle Cardiaque 2Document34 pagesPhysiologie Du Mucle Cardiaque 2Marcus MilitoPas encore d'évaluation
- SVT MatheoDocument8 pagesSVT Matheolionel rigaudPas encore d'évaluation
- 21 Tspe M1 FicheDocument2 pages21 Tspe M1 FicheMathilde JaeckelPas encore d'évaluation
- BIO 111 CHAP IXb 2021Document12 pagesBIO 111 CHAP IXb 2021meteweflorindaPas encore d'évaluation
- Le Muscle 1Document3 pagesLe Muscle 1travelwithoutborders2022Pas encore d'évaluation
- Système Musculaire 6-2 PDFDocument6 pagesSystème Musculaire 6-2 PDFLavi WilcalPas encore d'évaluation
- Cahierdetexte Chap3 TS SpeDocument3 pagesCahierdetexte Chap3 TS SpeFadhel SlamaPas encore d'évaluation
- Le Muscle 3Document3 pagesLe Muscle 3travelwithoutborders2022Pas encore d'évaluation
- CorrigeDocument2 pagesCorrigethiescoyabaPas encore d'évaluation
- Muscle 3Document3 pagesMuscle 3hkuuiPas encore d'évaluation
- Tissu Musculaire StriéDocument4 pagesTissu Musculaire StriéRayene HadibyPas encore d'évaluation
- MOTRICITE Par 2 DiapDocument79 pagesMOTRICITE Par 2 DiapRaphaël BikoahPas encore d'évaluation
- CCF Chap11 HCN ManiocDocument5 pagesCCF Chap11 HCN Maniocloris.yohanPas encore d'évaluation
- CM Physiologie MusculaireDocument20 pagesCM Physiologie Musculairekouirass.mohamedPas encore d'évaluation
- Physio Du MuscleDocument19 pagesPhysio Du Musclesamcruz31100% (1)
- Physiologie Du MuscleDocument33 pagesPhysiologie Du MuscleYannick NdriPas encore d'évaluation
- Receuil Physio CardiaqueDocument7 pagesReceuil Physio CardiaqueDavs PhobaPas encore d'évaluation
- 22 - Electrophysiologie de La Cellule Myocardique MDocument3 pages22 - Electrophysiologie de La Cellule Myocardique MAmira Djihene El MecherfiPas encore d'évaluation
- Le Cytosquelette RésuméDocument5 pagesLe Cytosquelette RésuméEva BrovedaniPas encore d'évaluation
- MOSTA Physiologie de La Jonction NeuromusculaireDocument27 pagesMOSTA Physiologie de La Jonction NeuromusculaireSamia BelabbasPas encore d'évaluation
- Physio2an Cardio-Notes Cours2018 PDFDocument48 pagesPhysio2an Cardio-Notes Cours2018 PDFOscar Homme de L'ombrePas encore d'évaluation
- Physiopathologie Des Troubles de RythmeDocument9 pagesPhysiopathologie Des Troubles de Rythmekarim mounaouirPas encore d'évaluation
- Le Cytosquelette1Document18 pagesLe Cytosquelette1Radjaa BerroubaPas encore d'évaluation
- L Utilisation de L ATP Par La Fibre Musculaire PDFDocument27 pagesL Utilisation de L ATP Par La Fibre Musculaire PDFsambaPas encore d'évaluation
- Acides nucléiques: Les Grands Articles d'UniversalisD'EverandAcides nucléiques: Les Grands Articles d'UniversalisPas encore d'évaluation
- Surrénales: Les Grands Articles d'UniversalisD'EverandSurrénales: Les Grands Articles d'UniversalisPas encore d'évaluation
- Plan Étude PotionsDocument63 pagesPlan Étude PotionsMelPas encore d'évaluation
- ABC de La ChiromancieDocument47 pagesABC de La ChiromancieToky Andrianintsoa100% (1)
- TD CodageDocument2 pagesTD CodageTeuma Mbezi MichelPas encore d'évaluation
- Grammatik13 Lieber BesserDocument2 pagesGrammatik13 Lieber BesserLasigo MokafiPas encore d'évaluation
- Science Vie Guerre R HistoireDocument132 pagesScience Vie Guerre R HistoireFlavio JúniorPas encore d'évaluation
- Bohemien en VoyageDocument6 pagesBohemien en VoyageNeilPas encore d'évaluation
- Cna CanDocument5 pagesCna CanAnass GmiraPas encore d'évaluation
- MDT218AJ10ENC25Document8 pagesMDT218AJ10ENC25mordidomiPas encore d'évaluation
- Les Lesions Elem PolycoDocument3 pagesLes Lesions Elem PolycoLarissa CompaorePas encore d'évaluation
- Activité 1: Les Médias Et La DémocratieDocument2 pagesActivité 1: Les Médias Et La DémocratielolaPas encore d'évaluation
- La Recherche 2022-10 pp138-139Document2 pagesLa Recherche 2022-10 pp138-139Virgule de GuillemetsPas encore d'évaluation
- FSSPX-Rome - Appel Du Superieur Du District D'asieDocument3 pagesFSSPX-Rome - Appel Du Superieur Du District D'asieCaveNeCadasPas encore d'évaluation
- Adjectif Verbal Et Participe Présent en FrançaisDocument37 pagesAdjectif Verbal Et Participe Présent en Françaisyoulmafia99Pas encore d'évaluation
- Généralités Com PharmacieDocument37 pagesGénéralités Com PharmacieMOHAMEDTAHA ZAHIRPas encore d'évaluation
- Chapitre 1 Analyse DescriptiveDocument4 pagesChapitre 1 Analyse Descriptivejubaar14Pas encore d'évaluation
- BSG 1erDocument64 pagesBSG 1erAbdoulaye Djibril BabaPas encore d'évaluation
- Manuel Du Pilote Genie de La Reaction1Document33 pagesManuel Du Pilote Genie de La Reaction1RandaPas encore d'évaluation
- 6 7271 4e59374d PDFDocument25 pages6 7271 4e59374d PDFSAEC LIBERTEPas encore d'évaluation
- GrammaireDocument12 pagesGrammaireAnmol DhillonPas encore d'évaluation
- MandarineCE2 Guide Pedagogique Chapitre01Document19 pagesMandarineCE2 Guide Pedagogique Chapitre01Camille CousinPas encore d'évaluation
- GRILLE D AUDIT PostanalytiqueDocument11 pagesGRILLE D AUDIT PostanalytiqueSKPACAPas encore d'évaluation
- Vertiges 2013 08 10 2013Document69 pagesVertiges 2013 08 10 2013Abdelmoumen DjazairiPas encore d'évaluation
- Les Pronoms en Et Y - Grammaire Delf A2Document4 pagesLes Pronoms en Et Y - Grammaire Delf A2Mondher DhahriPas encore d'évaluation
- Myopathies Inflammatoires Idiopathiques DMDocument8 pagesMyopathies Inflammatoires Idiopathiques DMTtPas encore d'évaluation
- Le Strabisme Cours Magistral FinalDocument59 pagesLe Strabisme Cours Magistral FinalNassim OuabouPas encore d'évaluation