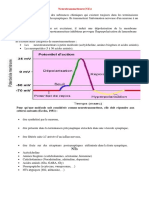
Vous aimerez peut-être aussi
- NEUROPHYSIOLOGIEDocument47 pagesNEUROPHYSIOLOGIEPrince AlegaPas encore d'évaluation
- Physiologie LA JONCTION NEUROMUSCULAIREDocument9 pagesPhysiologie LA JONCTION NEUROMUSCULAIREtoni.slmPas encore d'évaluation
- Electro - Syst Nerveux - CHAP IV B Transmission NeuromusculaireDocument68 pagesElectro - Syst Nerveux - CHAP IV B Transmission NeuromusculaireAliou LabaykaPas encore d'évaluation
- Activité MsculaireDocument7 pagesActivité MsculaireameniPas encore d'évaluation
- MOSTA Physiologie de La Jonction NeuromusculaireDocument27 pagesMOSTA Physiologie de La Jonction NeuromusculaireSamia BelabbasPas encore d'évaluation
- Synapse Et Transmission SynaptiqueDocument10 pagesSynapse Et Transmission SynaptiqueFaculté De Médecine BécharPas encore d'évaluation
- 6 - Transmission SynaptiqueDocument7 pages6 - Transmission Synaptiquemintchimechak2Pas encore d'évaluation
- 2-Physiologie MusculaireDocument44 pages2-Physiologie MusculaireIslęm OuaribPas encore d'évaluation
- 08-Physiologie de La Jonction NeuromusculaireDocument26 pages08-Physiologie de La Jonction NeuromusculaireBook HunterPas encore d'évaluation
- 02 Generalites Sur Les CuraresDocument12 pages02 Generalites Sur Les CuraresMokrane AmmourPas encore d'évaluation
- P Motricité (C. Quairiaux) - 26.03.21Document5 pagesP Motricité (C. Quairiaux) - 26.03.21nasten16fhPas encore d'évaluation
- Chapitre4.1 Biochimie Cellulaire Et FonctionnelleDocument7 pagesChapitre4.1 Biochimie Cellulaire Et Fonctionnelledraboissouf174Pas encore d'évaluation
- 03 - 2 - Proprietes Mecaniques Du Muscle - ContractionDocument30 pages03 - 2 - Proprietes Mecaniques Du Muscle - ContractionGloria KitonPas encore d'évaluation
- Support de Cours Physiologie Et Physiopathologie de La Cellule Musculaire ABED NousseibaDocument33 pagesSupport de Cours Physiologie Et Physiopathologie de La Cellule Musculaire ABED NousseibaSamwil SUANONPas encore d'évaluation
- Le Fonctionnement Du Muscle Squelettique: Magazine de Sciences de La Vie Et de La TerreDocument28 pagesLe Fonctionnement Du Muscle Squelettique: Magazine de Sciences de La Vie Et de La TerreSifddin GuidPas encore d'évaluation
- Physique Et TéléphoniqueDocument4 pagesPhysique Et TéléphoniquemeyemobiangjPas encore d'évaluation
- CorrigeDocument2 pagesCorrigethiescoyabaPas encore d'évaluation
- Role Des Jonctions Neuromusculaires 0Document3 pagesRole Des Jonctions Neuromusculaires 0modiop135Pas encore d'évaluation
- SN TP2 Synapse Correction PDFDocument5 pagesSN TP2 Synapse Correction PDFMankeur GueyePas encore d'évaluation
- SEQ 4-COURS Redige-Comportement-ReflexeDocument6 pagesSEQ 4-COURS Redige-Comportement-ReflexeLadetPas encore d'évaluation
- Exo Propriete Tissu Nerveux 7dDocument25 pagesExo Propriete Tissu Nerveux 7dNiangPas encore d'évaluation
- La Plaque MotriceDocument22 pagesLa Plaque MotriceTourgout Alb CengizPas encore d'évaluation
- 14 Histophysiologie Du Tissu NerveuxDocument7 pages14 Histophysiologie Du Tissu NerveuxGeorge BorsPas encore d'évaluation
- Cours Mucle RénovéDocument11 pagesCours Mucle Rénovémohamed.aziz.amri.11Pas encore d'évaluation
- Cours 12 - La Jonction NeuromusculaireDocument1 pageCours 12 - La Jonction NeuromusculaireSandra GomesPas encore d'évaluation
- 19a Transmis SynaptDocument15 pages19a Transmis SynaptTalel TouaitiPas encore d'évaluation
- Chapitre 9 - Réflexe Myotatique Et Message NerveuxDocument10 pagesChapitre 9 - Réflexe Myotatique Et Message NerveuxYousra INDIAPas encore d'évaluation
- Les Neurotransmetteurs PacesDocument30 pagesLes Neurotransmetteurs PacesValentin GeorgetPas encore d'évaluation
- ch10 Solutionnaire 1569538426Document12 pagesch10 Solutionnaire 1569538426Wilfried KouaoPas encore d'évaluation
- 4-La Transmission Synaptique PDFDocument14 pages4-La Transmission Synaptique PDFNara ĪntiSsãrPas encore d'évaluation
- Séance N°4 - : TUTORAT UE Physiologie 2012-2013Document4 pagesSéance N°4 - : TUTORAT UE Physiologie 2012-2013fati imadPas encore d'évaluation
- Neuro 2Document12 pagesNeuro 2tinaPas encore d'évaluation
- La Contraction MusculaireDocument6 pagesLa Contraction MusculaireLim BunkimPas encore d'évaluation
- La Contraction Musculaire PDFDocument9 pagesLa Contraction Musculaire PDFFaculté De Médecine Béchar50% (2)
- Le Muscle SquelettiqueDocument4 pagesLe Muscle SquelettiqueGabriela TituPas encore d'évaluation
- ExocytoseDocument18 pagesExocytosecoursenligne.lille1100% (1)
- Les Muscles LissesDocument5 pagesLes Muscles LissesHassanElhassanPas encore d'évaluation
- Physiologie Du Muscle Et de La Contraction MusculaireDocument5 pagesPhysiologie Du Muscle Et de La Contraction MusculaireImane BenderazPas encore d'évaluation
- Groupe 3Document13 pagesGroupe 3ngueguimlaurellaPas encore d'évaluation
- Physiologie Du Mucle Cardiaque 2Document34 pagesPhysiologie Du Mucle Cardiaque 2Marcus MilitoPas encore d'évaluation
- Les ReflexesDocument2 pagesLes ReflexeslozayPas encore d'évaluation
- Chapitre Iv Les Cellules ExcitablesDocument77 pagesChapitre Iv Les Cellules ExcitablesSARA IBRAHIMIPas encore d'évaluation
- Tissus MusculairesDocument16 pagesTissus MusculairesandriePas encore d'évaluation
- Electrophysiologie CardiaqueT PDFDocument3 pagesElectrophysiologie CardiaqueT PDFnadaPas encore d'évaluation
- La Cellule Nerveuse W.PDF KKDocument6 pagesLa Cellule Nerveuse W.PDF KKas.cherfaouiPas encore d'évaluation
- Chapitre 3 Le Système NerveuxDocument26 pagesChapitre 3 Le Système NerveuxdfhohfeicxhPas encore d'évaluation
- Electrophysiologie CardiaqueDocument6 pagesElectrophysiologie Cardiaqueelmkhailiabdellah93Pas encore d'évaluation
- C6.La Transmission SynaptiqueDocument19 pagesC6.La Transmission SynaptiqueAbdelOuahidSenhadjiPas encore d'évaluation
- Les Tissus MusculairesDocument11 pagesLes Tissus MusculairesZoulkiffiPas encore d'évaluation
- Resume de NeurophysiologieDocument5 pagesResume de NeurophysiologieMoussa DIAFOPas encore d'évaluation
- La Transmission Synaptique 1Document6 pagesLa Transmission Synaptique 1safemindPas encore d'évaluation
- Khaiter Devoir Numero 1Document5 pagesKhaiter Devoir Numero 1Y2kMagiikPas encore d'évaluation
- Réflexe TspéDocument8 pagesRéflexe TspéambreletallecPas encore d'évaluation
- TD N°3 de Physiologie AnimaleDocument10 pagesTD N°3 de Physiologie AnimaleMamadou lamine DiattaPas encore d'évaluation
- 1 - Electrophysiologie Cardiaque-Converti-1Document43 pages1 - Electrophysiologie Cardiaque-Converti-1ama23niPas encore d'évaluation
- Physio2an Cardio-Notes Cours2018 PDFDocument48 pagesPhysio2an Cardio-Notes Cours2018 PDFOscar Homme de L'ombrePas encore d'évaluation
- 5 - Physiologie Du NeuroneDocument8 pages5 - Physiologie Du Neuronemintchimechak2Pas encore d'évaluation
- Cahierdetexte Chap3 TS SpeDocument3 pagesCahierdetexte Chap3 TS SpeFadhel SlamaPas encore d'évaluation
- 1.physiologie Cardiaque Tanger Cours PR El Hangouche Part 1Document104 pages1.physiologie Cardiaque Tanger Cours PR El Hangouche Part 1elbakkalilibrairiePas encore d'évaluation
- Neurobiologie: Les Grands Articles d'UniversalisD'EverandNeurobiologie: Les Grands Articles d'UniversalisPas encore d'évaluation
- Tdiettdii: Nadh Nadh - 1 - 1 Nadh - 1 - 1Document6 pagesTdiettdii: Nadh Nadh - 1 - 1 Nadh - 1 - 1youssefbgh03Pas encore d'évaluation
- Cours de Biologie Cellulaire Rabat 2017-2018Document98 pagesCours de Biologie Cellulaire Rabat 2017-2018Badr ZbidaPas encore d'évaluation
- Liens Et Dates Pour Les Questions Ue1Document1 pageLiens Et Dates Pour Les Questions Ue1ttahaben07Pas encore d'évaluation
- Biochimie StructuraleDocument169 pagesBiochimie StructuraleGHERMI .M100% (7)
- 618 Cas CholestaseDocument10 pages618 Cas Cholestasereflexionperdue56Pas encore d'évaluation
- 2.7 TD Biologie Mol Synthese Des Proteines P CuneyDocument7 pages2.7 TD Biologie Mol Synthese Des Proteines P CuneyslimdamakPas encore d'évaluation
- Structure de l'ADNDocument14 pagesStructure de l'ADNrafiq ahmed zaidPas encore d'évaluation
- CYCLE TRICARBOXYLIQUE DE KREBS Vers EtudDocument30 pagesCYCLE TRICARBOXYLIQUE DE KREBS Vers EtudLaira ImiPas encore d'évaluation
- Chapitre 4 Marqueurs de La RIDocument32 pagesChapitre 4 Marqueurs de La RIMona PleyersPas encore d'évaluation
- Biologie PDFDocument30 pagesBiologie PDFElysee AmoakonPas encore d'évaluation
- Voie Des Pentoses PhosphatesDocument26 pagesVoie Des Pentoses Phosphatescep100% (1)
- Cours Exploration Du Diabéte Ecole de SantéDocument87 pagesCours Exploration Du Diabéte Ecole de SantéJlizi MalekPas encore d'évaluation
- Thymus VilgarisDocument141 pagesThymus Vilgarisbouzina ibrahimPas encore d'évaluation
- Metabolisme Xeno IntroDocument12 pagesMetabolisme Xeno IntroOlaya MenaceurPas encore d'évaluation
- Bio InformatiqueDocument199 pagesBio Informatiquerobndoh100% (1)
- GENETIQUEDocument2 pagesGENETIQUEFéz EyPas encore d'évaluation
- 10 - Cycle Et Senescence Diff Et Mort Cellulaire Cours 2022Document92 pages10 - Cycle Et Senescence Diff Et Mort Cellulaire Cours 2022Thuy TranPas encore d'évaluation
- Génétique II Chap I SV-S5Document59 pagesGénétique II Chap I SV-S5Heedyyl MaatoufPas encore d'évaluation
- Génotoxicité Mutagénèse TératogenèseDocument10 pagesGénotoxicité Mutagénèse TératogenèseRaid YaSsinePas encore d'évaluation
- Chapitre IVDocument6 pagesChapitre IVØüssÂmāPas encore d'évaluation
- TD3 CorrigesDocument18 pagesTD3 CorrigesDjawed BoutPas encore d'évaluation
- Document 20Document2 pagesDocument 20nizar rhazaliPas encore d'évaluation
- TD.03. BiomolDocument43 pagesTD.03. BiomolChawki MokademPas encore d'évaluation
- 2015 KOMAS Narcisse PatriceDocument67 pages2015 KOMAS Narcisse PatriceMaaz NasimPas encore d'évaluation
- Thèse Aymen 2011Document221 pagesThèse Aymen 2011Aymen Ezzine100% (1)
- Physiologie Du SPORTDocument29 pagesPhysiologie Du SPORTAbdessamad NfissiPas encore d'évaluation
- Bilan Thermique Dune Chambre FroideDocument34 pagesBilan Thermique Dune Chambre FroidestifbensalehPas encore d'évaluation
- SVT 2PC SimiliDocument7 pagesSVT 2PC Similitouahri aymane100% (1)
- Socle Commun - 2A AgronomieDocument46 pagesSocle Commun - 2A AgronomieLina AdamPas encore d'évaluation
- TD2 BA 171 Partie A 2013 2014Document2 pagesTD2 BA 171 Partie A 2013 2014Berthe Aristide Ngue NkongoPas encore d'évaluation