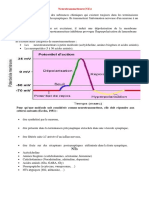
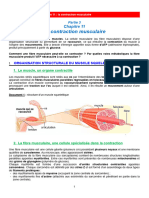
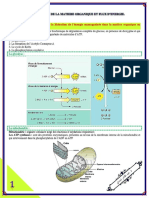
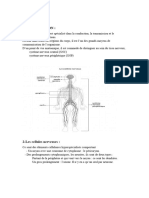
Vous aimerez peut-être aussi
- Physio1an31-7jonction NeuromusculaireDocument15 pagesPhysio1an31-7jonction Neuromusculaireelidrissimaryam733Pas encore d'évaluation
- DCEM1 - Pharmacologie - Chapitre 19 - Les Antinicotiniques - Septembre 2005 PDFDocument4 pagesDCEM1 - Pharmacologie - Chapitre 19 - Les Antinicotiniques - Septembre 2005 PDFPaul fatheadPas encore d'évaluation
- MOSTA Physiologie de La Jonction NeuromusculaireDocument27 pagesMOSTA Physiologie de La Jonction NeuromusculaireSamia BelabbasPas encore d'évaluation
- Physiologie LA JONCTION NEUROMUSCULAIREDocument9 pagesPhysiologie LA JONCTION NEUROMUSCULAIREtoni.slmPas encore d'évaluation
- 2-Physiologie MusculaireDocument44 pages2-Physiologie MusculaireIslęm OuaribPas encore d'évaluation
- PHARMACOLOGIE Des SystèmesDocument96 pagesPHARMACOLOGIE Des SystèmesRidou anePas encore d'évaluation
- Electro - Syst Nerveux - CHAP IV B Transmission NeuromusculaireDocument68 pagesElectro - Syst Nerveux - CHAP IV B Transmission NeuromusculaireAliou LabaykaPas encore d'évaluation
- NEUROPHYSIOLOGIEDocument47 pagesNEUROPHYSIOLOGIEPrince AlegaPas encore d'évaluation
- ch10 Solutionnaire 1569538426Document12 pagesch10 Solutionnaire 1569538426Wilfried KouaoPas encore d'évaluation
- 03 - 2 - Proprietes Mecaniques Du Muscle - ContractionDocument30 pages03 - 2 - Proprietes Mecaniques Du Muscle - ContractionGloria KitonPas encore d'évaluation
- Système Parasympathique 22-23Document16 pagesSystème Parasympathique 22-23MaryM SaidiPas encore d'évaluation
- Activité MsculaireDocument7 pagesActivité MsculaireameniPas encore d'évaluation
- La Contraction MusculaireDocument6 pagesLa Contraction MusculaireLim BunkimPas encore d'évaluation
- 4.3.2.mécanismes de La Contractio1Document4 pages4.3.2.mécanismes de La Contractio1Ayoub LakhdariPas encore d'évaluation
- Definitions SVT BiofDocument5 pagesDefinitions SVT BiofhadolaPas encore d'évaluation
- La Plaque MotriceDocument22 pagesLa Plaque MotriceTourgout Alb CengizPas encore d'évaluation
- 08-Physiologie de La Jonction NeuromusculaireDocument26 pages08-Physiologie de La Jonction NeuromusculaireBook HunterPas encore d'évaluation
- Khaiter Devoir Numero 1Document5 pagesKhaiter Devoir Numero 1Y2kMagiikPas encore d'évaluation
- Le Muscle NormalDocument41 pagesLe Muscle NormalHelena KenguePas encore d'évaluation
- AcetylcholineDocument9 pagesAcetylcholineSanaAjjPas encore d'évaluation
- Curarisants: Étapes de La CurarisationDocument9 pagesCurarisants: Étapes de La CurarisationAyoub LilouPas encore d'évaluation
- Définitions Des Termes Scientifiques (1) - 210220 - 191003Document12 pagesDéfinitions Des Termes Scientifiques (1) - 210220 - 191003Zakaria KRPas encore d'évaluation
- 14 Histophysiologie Du Tissu NerveuxDocument7 pages14 Histophysiologie Du Tissu NerveuxGeorge BorsPas encore d'évaluation
- Cours de Reeducation VestibulaireDocument106 pagesCours de Reeducation VestibulaireZOHEIR RAMDANE100% (1)
- Le Fonctionnement Du Muscle Squelettique: Magazine de Sciences de La Vie Et de La TerreDocument28 pagesLe Fonctionnement Du Muscle Squelettique: Magazine de Sciences de La Vie Et de La TerreSifddin GuidPas encore d'évaluation
- La Transmission Synaptique 1Document6 pagesLa Transmission Synaptique 1safemindPas encore d'évaluation
- Les Définitions SVTDocument20 pagesLes Définitions SVTYassmine Aj100% (2)
- Neuro 2Document12 pagesNeuro 2tinaPas encore d'évaluation
- Vocabulaires SVT 2BAC BIOF-1Document10 pagesVocabulaires SVT 2BAC BIOF-1Abderrahim Boulefra100% (1)
- Le Muscle SquelettiqueDocument4 pagesLe Muscle SquelettiqueGabriela TituPas encore d'évaluation
- Les Neurotransmetteurs PacesDocument30 pagesLes Neurotransmetteurs PacesValentin GeorgetPas encore d'évaluation
- Physiologie Bronchique CorrigéDocument18 pagesPhysiologie Bronchique CorrigéyecinePas encore d'évaluation
- La Contraction MusculaireDocument11 pagesLa Contraction MusculaireJihadi AbbouPas encore d'évaluation
- Chapitre 3 Le Système NerveuxDocument26 pagesChapitre 3 Le Système NerveuxdfhohfeicxhPas encore d'évaluation
- Définitions Des Termes Scientifiques Unité 1Document2 pagesDéfinitions Des Termes Scientifiques Unité 1assafarPas encore d'évaluation
- P Motricité (C. Quairiaux) - 26.03.21Document5 pagesP Motricité (C. Quairiaux) - 26.03.21nasten16fhPas encore d'évaluation
- Les Muscles LissesDocument5 pagesLes Muscles LissesHassanElhassanPas encore d'évaluation
- 5b4da16fd3e78 3Document33 pages5b4da16fd3e78 3Fabiola FatranPas encore d'évaluation
- Neuromusculaire - Les Points Essentiels M 4Document4 pagesNeuromusculaire - Les Points Essentiels M 4Guana TibiPas encore d'évaluation
- Exposé Biologie CellulaireDocument13 pagesExposé Biologie CellulaireRemadji vieriPas encore d'évaluation
- Physiologie Du Muscle Et de La Contraction MusculaireDocument5 pagesPhysiologie Du Muscle Et de La Contraction MusculaireImane BenderazPas encore d'évaluation
- Physiologie Du Mucle Cardiaque 2Document34 pagesPhysiologie Du Mucle Cardiaque 2Marcus MilitoPas encore d'évaluation
- Cours Des Anesthesiques (Locaux Et Généraux) 2022-2023Document15 pagesCours Des Anesthesiques (Locaux Et Généraux) 2022-2023Aissa Aymen BnPas encore d'évaluation
- Les ReflexesDocument2 pagesLes ReflexeslozayPas encore d'évaluation
- Cours Mucle RénovéDocument11 pagesCours Mucle Rénovémohamed.aziz.amri.11Pas encore d'évaluation
- Bilan Du Chapitre 11 - La Contraction MusculaireDocument12 pagesBilan Du Chapitre 11 - La Contraction MusculaireYousra INDIAPas encore d'évaluation
- Bestcours L'essentiel À Retenir PDFDocument22 pagesBestcours L'essentiel À Retenir PDFElamrani KarimPas encore d'évaluation
- Groupe 3Document13 pagesGroupe 3ngueguimlaurellaPas encore d'évaluation
- Photosynthèse: Les Grands Articles d'UniversalisD'EverandPhotosynthèse: Les Grands Articles d'UniversalisPas encore d'évaluation
- La Contraction Musculaire PDFDocument9 pagesLa Contraction Musculaire PDFFaculté De Médecine Béchar50% (2)
- Les Neuromediateurs 2024Document94 pagesLes Neuromediateurs 2024Corona CoronaPas encore d'évaluation
- Communication NerveuseDocument5 pagesCommunication NerveusemerzougPas encore d'évaluation
- Chapitre4.1 Biochimie Cellulaire Et FonctionnelleDocument7 pagesChapitre4.1 Biochimie Cellulaire Et Fonctionnelledraboissouf174Pas encore d'évaluation
- 3-Cours Endocrinologie Et Régulation Des Systèmes Chapitre 2 M1 TOXICOLOGIE 2020-2021 BOUBEKRIDocument32 pages3-Cours Endocrinologie Et Régulation Des Systèmes Chapitre 2 M1 TOXICOLOGIE 2020-2021 BOUBEKRIBERDJOUH YoucefPas encore d'évaluation
- Corrigé Muscles Métropole Sujet 1 2021Document8 pagesCorrigé Muscles Métropole Sujet 1 2021bellayerdianePas encore d'évaluation
- La Cellule Nerveuse W.PDF KKDocument6 pagesLa Cellule Nerveuse W.PDF KKas.cherfaouiPas encore d'évaluation
- Les Tissus MusculairesDocument11 pagesLes Tissus MusculairesZoulkiffiPas encore d'évaluation
- SVT MatheoDocument8 pagesSVT Matheolionel rigaudPas encore d'évaluation
- Electrophysiologie CardiaqueDocument6 pagesElectrophysiologie Cardiaqueelmkhailiabdellah93Pas encore d'évaluation
- Surrénales: Les Grands Articles d'UniversalisD'EverandSurrénales: Les Grands Articles d'UniversalisPas encore d'évaluation
- Allo-Immunisation RhesusDocument13 pagesAllo-Immunisation RhesusSekou NiangPas encore d'évaluation
- Exposé Ophtalmologie Groupe 05 FinalDocument23 pagesExposé Ophtalmologie Groupe 05 FinalRemadji vieriPas encore d'évaluation
- Glandes EndocrinesDocument1 pageGlandes EndocrinesDi BenamoPas encore d'évaluation
- 4-Anatomie de l'ŒSOPAHAGEDocument5 pages4-Anatomie de l'ŒSOPAHAGEpc lenovoPas encore d'évaluation
- Controle Systéme Nerveux Et MusculaireDocument2 pagesControle Systéme Nerveux Et MusculaireAziz AdlPas encore d'évaluation
- CoursDocument14 pagesCoursPHDIAPas encore d'évaluation
- Immuno 2Document20 pagesImmuno 2TALHAPas encore d'évaluation
- Les Reponses Immunitaires Cours MaDocument15 pagesLes Reponses Immunitaires Cours Mahoussame ben el hafianePas encore d'évaluation
- Poly Meth0501 TP1 ImmunohistochimieDocument6 pagesPoly Meth0501 TP1 ImmunohistochimieAnonymous mSKAaRUyydPas encore d'évaluation
- Embryogenèse SomatiqueDocument20 pagesEmbryogenèse SomatiqueAyoub KhouyaniPas encore d'évaluation
- Amenorhées PrimitivesDocument33 pagesAmenorhées PrimitivesZAKARIA ZAOUIAPas encore d'évaluation
- Cours Boucherie Academy Des Chefs 2 Zouhaier ZouariDocument62 pagesCours Boucherie Academy Des Chefs 2 Zouhaier ZouariSalma Ben HalimaPas encore d'évaluation
- HématologieDocument23 pagesHématologieCorona CoronaPas encore d'évaluation
- 03 - La ColorationDocument40 pages03 - La ColorationNouhaila YahdihPas encore d'évaluation
- Diue2015m1 Aube PancreasDocument64 pagesDiue2015m1 Aube PancreasMeriem SidPas encore d'évaluation
- Physiologie de L'oreilleDocument31 pagesPhysiologie de L'oreilleMonde De CharlottePas encore d'évaluation
- SN Chap3 Transmission SynaptiqueDocument7 pagesSN Chap3 Transmission SynaptiqueKevin HeimanuPas encore d'évaluation
- Scrotum Et TesticuleDocument36 pagesScrotum Et Testiculevaryvira6677Pas encore d'évaluation
- Anatomie QCMDocument11 pagesAnatomie QCMSALMAN KHAYATIPas encore d'évaluation
- E1 Sujet Bts DietDocument10 pagesE1 Sujet Bts DietBrahimKhomsiPas encore d'évaluation
- 10 - Pancréatite Aiguë - Google Docs..Document8 pages10 - Pancréatite Aiguë - Google Docs..aicha aPas encore d'évaluation
- 21 Tspe M1 FicheDocument2 pages21 Tspe M1 FicheMathilde JaeckelPas encore d'évaluation
- Chapitre 1 SangDocument43 pagesChapitre 1 SangNorhaine Nina100% (1)
- Produits Haoqi 1 1Document48 pagesProduits Haoqi 1 1mariéme senePas encore d'évaluation
- ANNAMED HistologieDocument48 pagesANNAMED Histologieyahya bakhriPas encore d'évaluation
- Les Appareils FonctionnelsDocument35 pagesLes Appareils FonctionnelsabdennebiyPas encore d'évaluation
- Hypertrophies Cavitaires - Dr. AbbouDocument4 pagesHypertrophies Cavitaires - Dr. Abboukarima karimaPas encore d'évaluation
- Medecine Chinoise: Les OrganesDocument2 pagesMedecine Chinoise: Les OrganesJules HGPas encore d'évaluation
- Bac Science NatuDocument79 pagesBac Science NatuLine Hustling DollarPas encore d'évaluation
- Qe Residanat InternatDocument11 pagesQe Residanat InternatOumayma OutmaniPas encore d'évaluation