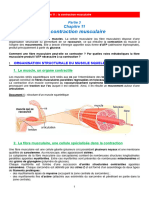
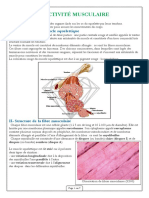
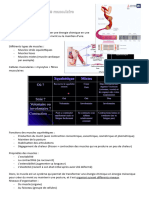
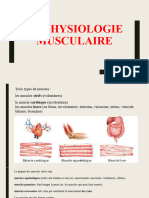
Vous aimerez peut-être aussi
- Definitions SVT BiofDocument5 pagesDefinitions SVT BiofhadolaPas encore d'évaluation
- Les Définitions SVTDocument20 pagesLes Définitions SVTYassmine Aj100% (1)
- Bilan Du Chapitre 11 - La Contraction MusculaireDocument12 pagesBilan Du Chapitre 11 - La Contraction MusculaireYousra INDIAPas encore d'évaluation
- Chapitre X Les MitochondriesDocument12 pagesChapitre X Les MitochondriesMichael Alafakis100% (1)
- Mito WRDocument3 pagesMito WRAbir M'ghariPas encore d'évaluation
- Activité MsculaireDocument7 pagesActivité MsculaireameniPas encore d'évaluation
- Résumé Chapitre 8Document2 pagesRésumé Chapitre 8k0k0sssit0Pas encore d'évaluation
- La Contraction Musculaire PDFDocument9 pagesLa Contraction Musculaire PDFFaculté De Médecine Béchar50% (2)
- Corrigé Muscles Métropole Sujet 1 2021Document8 pagesCorrigé Muscles Métropole Sujet 1 2021bellayerdianePas encore d'évaluation
- Cours de Tle D - Séquence 2Document4 pagesCours de Tle D - Séquence 2Tite DembelePas encore d'évaluation
- Définitions Des Termes Scientifiques Unité 1Document2 pagesDéfinitions Des Termes Scientifiques Unité 1assafarPas encore d'évaluation
- 2-Physiologie MusculaireDocument44 pages2-Physiologie MusculaireIslęm OuaribPas encore d'évaluation
- Définitions Des Termes Scientifiques (1) - 210220 - 191003Document12 pagesDéfinitions Des Termes Scientifiques (1) - 210220 - 191003Zakaria KRPas encore d'évaluation
- Origine de l'ATP Dans La Cellule Musculaire - Assistance Scolaire Personnalisée Et Gratuite - ASPDocument6 pagesOrigine de l'ATP Dans La Cellule Musculaire - Assistance Scolaire Personnalisée Et Gratuite - ASPYara ChehadePas encore d'évaluation
- ch10 Solutionnaire 1569538426Document12 pagesch10 Solutionnaire 1569538426Wilfried KouaoPas encore d'évaluation
- Physiologie Du Muscle Et de La Contraction MusculaireDocument5 pagesPhysiologie Du Muscle Et de La Contraction MusculaireImane BenderazPas encore d'évaluation
- Cahierdetexte Chap3 TS SpeDocument3 pagesCahierdetexte Chap3 TS SpeFadhel SlamaPas encore d'évaluation
- Les Tissus MusculairesDocument11 pagesLes Tissus MusculairesZoulkiffiPas encore d'évaluation
- Quelques Définitions Usuelles en SVT Option ProfDocument4 pagesQuelques Définitions Usuelles en SVT Option ProfAyoub Lilou0% (1)
- Khaiter Devoir Numero 1Document5 pagesKhaiter Devoir Numero 1Y2kMagiikPas encore d'évaluation
- Définitions 1 Libération de L Énergie Emmagasinée Dans La MaDocument3 pagesDéfinitions 1 Libération de L Énergie Emmagasinée Dans La MaYoussef AkrinePas encore d'évaluation
- FC1 GALUSCA Rappels Des Grandes Voies Metaboliques Et Leurs Integrations 10.01Document20 pagesFC1 GALUSCA Rappels Des Grandes Voies Metaboliques Et Leurs Integrations 10.01Romane chapuisPas encore d'évaluation
- La Contraction MusculaireDocument6 pagesLa Contraction MusculaireLim BunkimPas encore d'évaluation
- La Mitochondrie Est Un Organite Clos Possédant LesDocument32 pagesLa Mitochondrie Est Un Organite Clos Possédant LesFaresPas encore d'évaluation
- Les Muscles LissesDocument5 pagesLes Muscles LissesHassanElhassanPas encore d'évaluation
- Le Muscle SquelettiqueDocument4 pagesLe Muscle SquelettiqueGabriela TituPas encore d'évaluation
- Révision Spécialité SVT PDFDocument11 pagesRévision Spécialité SVT PDFGilles OleaPas encore d'évaluation
- CM Physiologie MusculaireDocument20 pagesCM Physiologie Musculairekouirass.mohamedPas encore d'évaluation
- La Contraction MusculaireDocument11 pagesLa Contraction MusculaireJihadi AbbouPas encore d'évaluation
- Physiologie Muscles Striés SquelettiquesDocument16 pagesPhysiologie Muscles Striés SquelettiquessafemindPas encore d'évaluation
- Vocabulaires SVT 2BAC BIOF-1Document10 pagesVocabulaires SVT 2BAC BIOF-1Abderrahim Boulefra100% (1)
- La Physiologie MusculaireDocument51 pagesLa Physiologie MusculaireJoronavalona RASAMIMANANAPas encore d'évaluation
- Chapitre4.1 Biochimie cellulaire et fonctionnelleDocument7 pagesChapitre4.1 Biochimie cellulaire et fonctionnelledraboissouf174Pas encore d'évaluation
- BILANS N°2 Chap 2-3-4 Thème 3Document5 pagesBILANS N°2 Chap 2-3-4 Thème 3TimPas encore d'évaluation
- 1.Physiologie musculaire 2023 (1)Document59 pages1.Physiologie musculaire 2023 (1)aminezebar3Pas encore d'évaluation
- Physiologie Du Mucle Cardiaque 2Document34 pagesPhysiologie Du Mucle Cardiaque 2Marcus MilitoPas encore d'évaluation
- UE1-Biochimie-9-Phosphorylation OxydativeDocument24 pagesUE1-Biochimie-9-Phosphorylation Oxydativesun-nee-chan9Pas encore d'évaluation
- UE2 - Biologie Cellulaire - Production D - Énergie Dans La CelluleDocument16 pagesUE2 - Biologie Cellulaire - Production D - Énergie Dans La Cellulesun-nee-chan9Pas encore d'évaluation
- La Contraction MusculaireDocument5 pagesLa Contraction MusculairekharratPas encore d'évaluation
- Les Mitochondries 1Document9 pagesLes Mitochondries 1Wissam TizaPas encore d'évaluation
- Exposé de Cytologie: ThèmeDocument41 pagesExposé de Cytologie: ThèmeFaresPas encore d'évaluation
- Chapitre 3 Maturation Des ProtéinesDocument34 pagesChapitre 3 Maturation Des Protéinesryadh2203Pas encore d'évaluation
- Biologie La CelluleDocument12 pagesBiologie La Cellule6z8nbjv76vPas encore d'évaluation
- Lettre IEPP n16 Mitochondrie v2 01 2016Document12 pagesLettre IEPP n16 Mitochondrie v2 01 2016manel yasminePas encore d'évaluation
- Contraction Musculaire CouleurDocument286 pagesContraction Musculaire Couleurmatis47215100% (1)
- Définition de ATPDocument2 pagesDéfinition de ATPmohbio_869948847Pas encore d'évaluation
- Production Et Role ATPDocument3 pagesProduction Et Role ATPWACDPas encore d'évaluation
- Physiologie Du MuscleDocument3 pagesPhysiologie Du MuscleTASMINE HAMZAPas encore d'évaluation
- Poly Histo 10-11 Tome2RANGUEIL PDFDocument94 pagesPoly Histo 10-11 Tome2RANGUEIL PDFmedPas encore d'évaluation
- Physiologie Vegetale 2 S4Document20 pagesPhysiologie Vegetale 2 S4hassan aitPas encore d'évaluation
- Support de Cours Physiologie Et Physiopathologie de La Cellule Musculaire ABED NousseibaDocument33 pagesSupport de Cours Physiologie Et Physiopathologie de La Cellule Musculaire ABED NousseibaSamwil SUANONPas encore d'évaluation
- FrançaisDocument5 pagesFrançaistiakola031Pas encore d'évaluation
- Résumé U-1 - P-2Document4 pagesRésumé U-1 - P-2ygtzdntp2j100% (1)
- Physio Du MuscleDocument19 pagesPhysio Du Musclesamcruz31100% (1)
- CM Muscle FRB 6 DiaDocument13 pagesCM Muscle FRB 6 DiaWilly Le DruillennecPas encore d'évaluation
- Correction - TD2 - BI - 125 - Structure - Et Fonction - de La - Cellule - AnimaleDocument5 pagesCorrection - TD2 - BI - 125 - Structure - Et Fonction - de La - Cellule - AnimaleBerthe Aristide Ngue NkongoPas encore d'évaluation
- UntitledDocument97 pagesUntitledGhita KhalilPas encore d'évaluation
- Photosynthèse: Les Grands Articles d'UniversalisD'EverandPhotosynthèse: Les Grands Articles d'UniversalisPas encore d'évaluation
- Acides nucléiques: Les Grands Articles d'UniversalisD'EverandAcides nucléiques: Les Grands Articles d'UniversalisPas encore d'évaluation
- Muscles: Les Grands Articles d'UniversalisD'EverandMuscles: Les Grands Articles d'UniversalisPas encore d'évaluation
- Casse NoisetteDocument13 pagesCasse Noisettelionel rigaudPas encore d'évaluation
- Lettre de MotivationDocument2 pagesLettre de Motivationlionel rigaudPas encore d'évaluation
- Bio George HistoireDocument2 pagesBio George Histoirelionel rigaudPas encore d'évaluation
- ArtDocument1 pageArtlionel rigaudPas encore d'évaluation
- PR Paration DELF B1 Hachette PDFDocument17 pagesPR Paration DELF B1 Hachette PDFVireak DyPas encore d'évaluation
- Ferment EursDocument98 pagesFerment Eursryma bouldjedjPas encore d'évaluation
- Q.C.M.génétique - Med 23 + PDFDocument7 pagesQ.C.M.génétique - Med 23 + PDFassssss100% (1)
- Myologie Membre InférieurDocument15 pagesMyologie Membre Inférieurbaptistechabance1Pas encore d'évaluation
- Physiologie de La LactationDocument18 pagesPhysiologie de La LactationÉli TePas encore d'évaluation
- Anatomie Du MaxillaireDocument15 pagesAnatomie Du MaxillaireAdina PopaPas encore d'évaluation
- Dix Exercices de Mise À La Terre Qui Amélioreront Votre VieDocument6 pagesDix Exercices de Mise À La Terre Qui Amélioreront Votre VieJohanne AnctilPas encore d'évaluation
- T3A2 Acti2 FCTT OvairesDocument4 pagesT3A2 Acti2 FCTT Ovairesbianca fPas encore d'évaluation
- Tumeurs Hypophysaires (Pr. Kesri)Document47 pagesTumeurs Hypophysaires (Pr. Kesri)Elixir'sPas encore d'évaluation
- Anapath L'essentielDocument5 pagesAnapath L'essentielTakishy MindLordPas encore d'évaluation
- BAC3 - Anatomie TopographiqueDocument344 pagesBAC3 - Anatomie TopographiqueLéa Winer100% (1)
- La VisionDocument3 pagesLa VisionFellag SaraPas encore d'évaluation
- Myologie Du Membre SupérieurDocument113 pagesMyologie Du Membre SupérieurAnwar El AraichiPas encore d'évaluation
- PHYSIOCARDIAQUEDocument41 pagesPHYSIOCARDIAQUEAnou KalengaPas encore d'évaluation
- Anatomie Du Système EndocrinienDocument44 pagesAnatomie Du Système EndocrinienvinickPas encore d'évaluation
- Cours Racines PDFDocument8 pagesCours Racines PDFSofiane BousifePas encore d'évaluation
- 1 Introduction A La Physiologie Des Glandes EndocrinesDocument16 pages1 Introduction A La Physiologie Des Glandes EndocrinesAlina Topor Psi-MedicalPas encore d'évaluation
- Paper 1 May 2021 Tz2Document22 pagesPaper 1 May 2021 Tz2fatenPas encore d'évaluation
- TD2 - RMDocument3 pagesTD2 - RMFatïma ZohraPas encore d'évaluation
- Recueil Dexercice Corriges en CMDocument89 pagesRecueil Dexercice Corriges en CMamine100% (3)
- Partie 1 Enzymo FEV 12Document61 pagesPartie 1 Enzymo FEV 12Limoune AliPas encore d'évaluation
- SOMESTHESIE-Document12 pagesSOMESTHESIE-mbouillesissoko55Pas encore d'évaluation
- GPI124Document1 pageGPI124sgyrdPas encore d'évaluation
- Anatomie Du Nez Et Des Sinus de La FaceDocument15 pagesAnatomie Du Nez Et Des Sinus de La FaceMarevaPas encore d'évaluation
- TD N°3Document3 pagesTD N°3ESSOME ESSOME OLIVIER STEPHANEPas encore d'évaluation