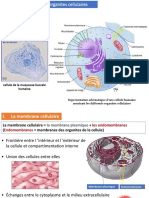
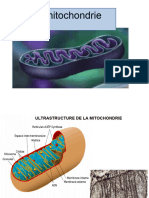
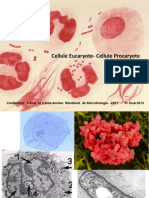
Vous aimerez peut-être aussi
- Membranes cellulaires: Les Grands Articles d'UniversalisD'EverandMembranes cellulaires: Les Grands Articles d'UniversalisPas encore d'évaluation
- Astus... Livre Tle D 2014 F-1-1Document168 pagesAstus... Livre Tle D 2014 F-1-1Rachid Dxpe93% (42)
- TD7 Les Organites de La Cellule PDFDocument42 pagesTD7 Les Organites de La Cellule PDFZineb DahmaniPas encore d'évaluation
- 1-Cours D IntroductionDocument40 pages1-Cours D Introductionml,zml,zPas encore d'évaluation
- La CelluleDocument6 pagesLa CelluleHossam AnaPas encore d'évaluation
- Biologie Cellulaire Et Biochimie Structural 2Document14 pagesBiologie Cellulaire Et Biochimie Structural 2bastaouiiPas encore d'évaluation
- Chapitre I Structure de La CelluleDocument107 pagesChapitre I Structure de La CelluleDescartes KpannePas encore d'évaluation
- AntifongiquesDocument21 pagesAntifongiquesyousra guendouzPas encore d'évaluation
- Biologie CellulaireDocument23 pagesBiologie CellulaireKévin Colin100% (3)
- BCHM1215Document51 pagesBCHM1215Asma ArfaouiPas encore d'évaluation
- Récepteurs MembranairesDocument56 pagesRécepteurs MembranairesAnonymous MKSfyYyODPPas encore d'évaluation
- 1.la MitochondrieDocument39 pages1.la MitochondrieAB AbPas encore d'évaluation
- La MitochondrieDocument7 pagesLa Mitochondrieilsane dawaleh geudiPas encore d'évaluation
- La Mitochondrie Est Un Organite Clos Possédant LesDocument32 pagesLa Mitochondrie Est Un Organite Clos Possédant LesFaresPas encore d'évaluation
- Les Mitochondries 1Document9 pagesLes Mitochondries 1Wissam TizaPas encore d'évaluation
- ChondriomeDocument25 pagesChondriomeYlinka SamiraPas encore d'évaluation
- 4 - MitochondriesDocument7 pages4 - MitochondriesVictor AllardPas encore d'évaluation
- UE2 - Biologie Cellulaire - Production D - Énergie Dans La CelluleDocument16 pagesUE2 - Biologie Cellulaire - Production D - Énergie Dans La Cellulesun-nee-chan9Pas encore d'évaluation
- Les MitochondriesDocument40 pagesLes MitochondriesRuth KoudPas encore d'évaluation
- 2022 MTC - DR MeradDocument8 pages2022 MTC - DR Meradkhouloudnemmouchi240721Pas encore d'évaluation
- La Mitochondrie - PremierePartieDocument11 pagesLa Mitochondrie - PremierePartieMoussa DIAFOPas encore d'évaluation
- Les Mitochondries Et Les ChloroplastesDocument20 pagesLes Mitochondries Et Les ChloroplastesMbaye AbouPas encore d'évaluation
- 9 MitochondriesDocument42 pages9 MitochondriesTyl HyuPas encore d'évaluation
- La Mitochondrie - RésuméDocument3 pagesLa Mitochondrie - RésuméEva BrovedaniPas encore d'évaluation
- Mitochondrie - WikipédiaDocument56 pagesMitochondrie - Wikipédia4xxstan2022Pas encore d'évaluation
- Production D'energie PDFDocument10 pagesProduction D'energie PDFSoubere MahamoudPas encore d'évaluation
- Cyto1an Mitochondrie2022aouatiDocument12 pagesCyto1an Mitochondrie2022aouatiAy ManPas encore d'évaluation
- Structure BactérienneDocument72 pagesStructure BactériennekhouildiPas encore d'évaluation
- Mito WRDocument3 pagesMito WRAbir M'ghariPas encore d'évaluation
- 5 MitochondrieDocument10 pages5 MitochondrieSõu ÂdPas encore d'évaluation
- Bcis 3Document25 pagesBcis 3rimas23rimas14Pas encore d'évaluation
- Lettre IEPP n16 Mitochondrie v2 01 2016Document12 pagesLettre IEPP n16 Mitochondrie v2 01 2016manel yasminePas encore d'évaluation
- Membrane PlasmiqueDocument7 pagesMembrane PlasmiqueSara BoudjemaiPas encore d'évaluation
- CytologieDocument43 pagesCytologieMouats Badreddine100% (2)
- Chap 2 MPDocument7 pagesChap 2 MPdramohandlhadjPas encore d'évaluation
- Anatomie 1 (Récupération Automatique)Document3 pagesAnatomie 1 (Récupération Automatique)Hossam AnaPas encore d'évaluation
- Bio CellulaireDocument44 pagesBio CellulaireTouri Na100% (1)
- 08.10.2015 Organisation Du Trafic IntracellulaireDocument17 pages08.10.2015 Organisation Du Trafic IntracellulaireEaPas encore d'évaluation
- Le Chondriome2021 CH8Document99 pagesLe Chondriome2021 CH8paul destinPas encore d'évaluation
- Sujeujet Bio Celle Session 2 PCEM1Document4 pagesSujeujet Bio Celle Session 2 PCEM1medPas encore d'évaluation
- Rachidi Walid p02Document59 pagesRachidi Walid p02Yasine McaPas encore d'évaluation
- 2021 - Cours La Cellule Version ComplèteDocument10 pages2021 - Cours La Cellule Version Complèteagathe surusPas encore d'évaluation
- BIO 111 - CHAP - VI - VII - Et - VIIIDocument9 pagesBIO 111 - CHAP - VI - VII - Et - VIIIBerthe Aristide Ngue NkongoPas encore d'évaluation
- Chapitre 5 Organite Semi Autonomes (Mitochondrie Chloroplaste) 2020 21Document52 pagesChapitre 5 Organite Semi Autonomes (Mitochondrie Chloroplaste) 2020 21cilekkoksuPas encore d'évaluation
- Cours de Cytologie, Embryologie Et Genetique Medicale by MBAILASSEM MBAINAIDA GEORGESDocument102 pagesCours de Cytologie, Embryologie Et Genetique Medicale by MBAILASSEM MBAINAIDA GEORGESMãhąmãţ Hãbįb Mõųşşã ĄhmąţPas encore d'évaluation
- Les-Mitochondries - Compressed 1Document33 pagesLes-Mitochondries - Compressed 1marie dominique patricia kouassiPas encore d'évaluation
- Biologie La CelluleDocument12 pagesBiologie La Cellule6z8nbjv76vPas encore d'évaluation
- Mitochondrie La Divine BacterieDocument17 pagesMitochondrie La Divine BacterieMOSCADELLIPas encore d'évaluation
- EchangesDocument12 pagesEchangesAmina assala Hadj MohamedPas encore d'évaluation
- Chap - 6 - Les Compartiments IntracellulairesDocument4 pagesChap - 6 - Les Compartiments IntracellulairesMoeanau GrangerPas encore d'évaluation
- 3.schema General Des Cellules Procaryotes Et Eucaryotes 2021-2022Document56 pages3.schema General Des Cellules Procaryotes Et Eucaryotes 2021-2022Le BoyaïstePas encore d'évaluation
- Cours de Biologie Cellulaire Rabat 2017-2018Document98 pagesCours de Biologie Cellulaire Rabat 2017-2018Badr ZbidaPas encore d'évaluation
- Bio Cell - Part 4Document20 pagesBio Cell - Part 4Zineb hmt27Pas encore d'évaluation
- Cours BCG UFR2SDocument19 pagesCours BCG UFR2SSirandou DiaoPas encore d'évaluation
- Physiologie 1 Synthèse Chapitre 3Document14 pagesPhysiologie 1 Synthèse Chapitre 3pierre Garcia-DamienPas encore d'évaluation
- Biologie Générale HumaineDocument10 pagesBiologie Générale HumaineSarah CabieuPas encore d'évaluation
- Quelques Définitions Usuelles en SVT Option ProfDocument4 pagesQuelques Définitions Usuelles en SVT Option ProfAyoub Lilou0% (1)
- MITOCHONDRIOPATHIESDocument51 pagesMITOCHONDRIOPATHIESLOKO Maoudi PaulettePas encore d'évaluation
- Httpsstaff - Univ-Batna2.dzsitesdefaultfilesabdessemed Samirafileschapitre 11 PDFDocument12 pagesHttpsstaff - Univ-Batna2.dzsitesdefaultfilesabdessemed Samirafileschapitre 11 PDFariel.2mariemPas encore d'évaluation
- 1.cours Biologie CellulaireDocument63 pages1.cours Biologie Cellulaireبدر الذهابيPas encore d'évaluation
- La Cellule PDFDocument4 pagesLa Cellule PDFnehemie nwayPas encore d'évaluation
- Endosymbiose Des MitochondriesDocument6 pagesEndosymbiose Des MitochondriesAmar AliPas encore d'évaluation
- Chapitre 1Document5 pagesChapitre 1Fatïma ZohraPas encore d'évaluation
- Sida, Des Bases Pour ComprendreDocument202 pagesSida, Des Bases Pour ComprendrePatuwianPas encore d'évaluation
- Chapitre 2 Partie 1-3Document9 pagesChapitre 2 Partie 1-3khaldi aminaPas encore d'évaluation
- La Contraction MusculaireDocument6 pagesLa Contraction MusculaireLim BunkimPas encore d'évaluation
- Cours No 02-La CelluleDocument13 pagesCours No 02-La CelluleFeriel FerielPas encore d'évaluation
- TD 1 Biologie Cellulaire 2020Document3 pagesTD 1 Biologie Cellulaire 2020wissalabPas encore d'évaluation
- 04 - La Transmission SynaptiqueDocument13 pages04 - La Transmission SynaptiqueNahla TrodiPas encore d'évaluation
- 6-Le Metabolisme Du FerDocument38 pages6-Le Metabolisme Du Ferdieudonne dahngwaPas encore d'évaluation
- CytobiologieDocument17 pagesCytobiologiedjaouad bourouaisPas encore d'évaluation
- IV.2. Les PlastesDocument45 pagesIV.2. Les PlastesPaulin BILLAUDPas encore d'évaluation
- Structure Des Aliments-2-22Document21 pagesStructure Des Aliments-2-22blopPas encore d'évaluation
- TD Séance 2 Pour ÉtudiantsDocument12 pagesTD Séance 2 Pour ÉtudiantsokPas encore d'évaluation
- Bio Cell - Annales Corrigés 2019-2022Document150 pagesBio Cell - Annales Corrigés 2019-2022Nathan CohenPas encore d'évaluation
- Techniques Membranaires À Gradient de PressionDocument13 pagesTechniques Membranaires À Gradient de Pressionguiguimol100% (1)
- SALIVATION. Excrétion - Processus Par Lequel Le Produit de La Sécrétion D'une Glande Est Rejeté Hors de Celle-Ci Par Un Ou Des Canaux.Document24 pagesSALIVATION. Excrétion - Processus Par Lequel Le Produit de La Sécrétion D'une Glande Est Rejeté Hors de Celle-Ci Par Un Ou Des Canaux.Saad OuakiaaPas encore d'évaluation
- BC-Constituants Chimiques de La CelluleDocument83 pagesBC-Constituants Chimiques de La CelluleZaynab Abdeljawad100% (1)
- Neurone Ben AhmedDocument12 pagesNeurone Ben Ahmeddan ebenePas encore d'évaluation
- Chapitre I Cours 1Document20 pagesChapitre I Cours 1Abderahmane KmrPas encore d'évaluation
- Microbiologiegnrale DR TOUAITIADocument36 pagesMicrobiologiegnrale DR TOUAITIAlina LaPas encore d'évaluation
- BalDocument7 pagesBalFATIMA EZZAHRAE KHRIBICHPas encore d'évaluation
- Chapitre 5 Erythropoà à Se Et GR 3A PH UM6SS 2022 EL MALIKI (Enregistrement Automatique)Document26 pagesChapitre 5 Erythropoà à Se Et GR 3A PH UM6SS 2022 EL MALIKI (Enregistrement Automatique)Amine HakimiPas encore d'évaluation
- CytologieDocument7 pagesCytologiedorissePas encore d'évaluation
- Acc - Bases de La SignalisationDocument5 pagesAcc - Bases de La Signalisationchahdsouf086Pas encore d'évaluation
- Cour 1 Culture Des Cellules Animales PDFDocument11 pagesCour 1 Culture Des Cellules Animales PDFAya NouiouaPas encore d'évaluation
- 10 Anesthesiques3Document45 pages10 Anesthesiques3Ro FaidaPas encore d'évaluation
- Biochimie Structurale Des Lipides PDFDocument37 pagesBiochimie Structurale Des Lipides PDFDr.Zakaria MFTPas encore d'évaluation
- Ziani Techniques de Biologie Moléculaire L3 Biologie Moléculaire1Document56 pagesZiani Techniques de Biologie Moléculaire L3 Biologie Moléculaire1soualahPas encore d'évaluation