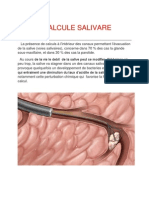
Télécharger au format pdf ou txt
Vous aimerez peut-être aussi
- Extraction Du Sucre de BetteraveDocument44 pagesExtraction Du Sucre de Betteraveabd100% (2)
- UE3, EC1 - Salivation - 18.09.2023Document7 pagesUE3, EC1 - Salivation - 18.09.2023okhan.kasimayPas encore d'évaluation
- Sécheresse BuccaleDocument26 pagesSécheresse BuccaleDt ImænëPas encore d'évaluation
- 5 - La Secheresse Buccale Et HypersialieDocument26 pages5 - La Secheresse Buccale Et HypersialieImane ABBASPas encore d'évaluation
- Salive 1Document45 pagesSalive 1Diane Stecy BIANG ENGONEPas encore d'évaluation
- Physiologie DigestiveDocument27 pagesPhysiologie Digestiveyaya camaraPas encore d'évaluation
- Physiologie Des Glandes SalivairesDocument5 pagesPhysiologie Des Glandes Salivairesasmaa.anime91Pas encore d'évaluation
- Les Glandes SalivairesDocument22 pagesLes Glandes SalivairesElmassal Sergeo100% (1)
- Etiopatogenia Cariei DentareDocument170 pagesEtiopatogenia Cariei DentareNelly AndriescuPas encore d'évaluation
- Introduction A La Pathologie Des Glandes SalivairesDocument7 pagesIntroduction A La Pathologie Des Glandes Salivaireslilly fritihPas encore d'évaluation
- La Salive: Réalisé ParDocument46 pagesLa Salive: Réalisé ParG kaoutherPas encore d'évaluation
- DigestionDocument3 pagesDigestionZaki ZakiPas encore d'évaluation
- Pathologie Des Glandes SalivairesDocument14 pagesPathologie Des Glandes SalivairesChouaib MeraoumiaPas encore d'évaluation
- Digestion Des Animaux D'élevageDocument31 pagesDigestion Des Animaux D'élevagePierre MonharoulPas encore d'évaluation
- Calcule SalivareDocument5 pagesCalcule SalivareEugenia MirosnicencoPas encore d'évaluation
- Lithiase SalivaireDocument18 pagesLithiase SalivaireDARGA AngePas encore d'évaluation
- 1-La Sécrétion SalivaireDocument4 pages1-La Sécrétion Salivairemedfarssi1029Pas encore d'évaluation
- Physiologie - Digestive ''''Document52 pagesPhysiologie - Digestive ''''Manassé MisengePas encore d'évaluation
- Physiologie DigestiveDocument49 pagesPhysiologie DigestiveAnou KalengaPas encore d'évaluation
- 1 - Physiologie de La Phase Bucco-OesophagienneDocument38 pages1 - Physiologie de La Phase Bucco-Oesophagienneroza jhjchvPas encore d'évaluation
- Odontlogie - Milieu BuccalDocument142 pagesOdontlogie - Milieu BuccalrajhasherPas encore d'évaluation
- Cours Paces Milieu Buccal - 2024Document161 pagesCours Paces Milieu Buccal - 2024Elior AkouriPas encore d'évaluation
- 2.phase Bucco-OesophagienneDocument10 pages2.phase Bucco-OesophagienneBer MouPas encore d'évaluation
- Capture D'écran . 2022-12-24 À 15.38.00Document46 pagesCapture D'écran . 2022-12-24 À 15.38.00Mangaka LightPas encore d'évaluation
- 2-Phase Bucco OesophagienneDocument7 pages2-Phase Bucco OesophagienneBer MouPas encore d'évaluation
- APP DIGESTIF DR DOUKKANEDocument28 pagesAPP DIGESTIF DR DOUKKANEamineje202Pas encore d'évaluation
- Cours 12 Digestif 2012Document41 pagesCours 12 Digestif 2012Ramona T. RosogaPas encore d'évaluation
- AZ Physio GastroDocument7 pagesAZ Physio GastroTinaPas encore d'évaluation
- Physiologie de La Mastication Et SalivationDocument7 pagesPhysiologie de La Mastication Et SalivationChaima benPas encore d'évaluation
- Les Glandes SalivairesDocument6 pagesLes Glandes SalivairesAdamou Djibo NénéPas encore d'évaluation
- Appareil Digestif 2ème Année Véto Cours 1 2021Document73 pagesAppareil Digestif 2ème Année Véto Cours 1 2021walid bb100% (1)
- 03-02 - 14h-15h - Bovine - Semio DigestiveDocument4 pages03-02 - 14h-15h - Bovine - Semio Digestivelibrairie MarhabaPas encore d'évaluation
- Les Glandes Salivaires - 231222 - 150743Document2 pagesLes Glandes Salivaires - 231222 - 150743aichamellouk20Pas encore d'évaluation
- La Déglutiton temps-WPS Office PDFDocument6 pagesLa Déglutiton temps-WPS Office PDFمصطفى كزوزPas encore d'évaluation
- Glandes SalivairesDocument1 pageGlandes SalivairesWalid BillalPas encore d'évaluation
- Physio Digest L1S2 Ziguinch 2022Document13 pagesPhysio Digest L1S2 Ziguinch 2022antaafall464Pas encore d'évaluation
- Digestion 1Document20 pagesDigestion 1douniaPas encore d'évaluation
- AUPIAIS 3Ã Me Annã e Bucco-Facial Oct 2022Document71 pagesAUPIAIS 3Ã Me Annã e Bucco-Facial Oct 2022doukkPas encore d'évaluation
- Cours - Appareil DigestifDocument19 pagesCours - Appareil Digestifiilhamita467Pas encore d'évaluation
- 3 - DéglutitionDocument17 pages3 - DéglutitionouazzanyPas encore d'évaluation
- DIGESTION Salive InsalivationDocument22 pagesDIGESTION Salive InsalivationMaroc Agro ConseilPas encore d'évaluation
- L'ulcère Gastro-DuodénaleDocument12 pagesL'ulcère Gastro-Duodénaleskeleton93Pas encore d'évaluation
- Cours D'anatomie Appareil-DigestifDocument11 pagesCours D'anatomie Appareil-DigestifACKA0% (1)
- Lithiase Des Glandes Salivaire 2023Document33 pagesLithiase Des Glandes Salivaire 2023Amadou sakhoPas encore d'évaluation
- Physio2an Digestive-Deglutition Motricite Oesophagienne2021abdelouahabDocument25 pagesPhysio2an Digestive-Deglutition Motricite Oesophagienne2021abdelouahabcdPas encore d'évaluation
- Les Fentes Vélo-PalatinesDocument6 pagesLes Fentes Vélo-Palatinesadelmiringui2Pas encore d'évaluation
- XérostomieDocument8 pagesXérostomieelghachi98Pas encore d'évaluation
- Anatomie Et Physiologie Du Systeme DigestifDocument49 pagesAnatomie Et Physiologie Du Systeme DigestifFerdinand Valère MebouinzPas encore d'évaluation
- Lithiases SalivairesDocument17 pagesLithiases SalivairesENT Doctor100% (2)
- EXPOSEDocument44 pagesEXPOSERania Prya MUTHUSAMY RENGASAMY100% (1)
- Physiologie de La Digestion 1ère Partie PDFDocument46 pagesPhysiologie de La Digestion 1ère Partie PDFBMA-medecine100% (2)
- 1-La Phase Bucco-Œsophagienne de La Digestion.1362058756Document56 pages1-La Phase Bucco-Œsophagienne de La Digestion.1362058756DëLux OwOPas encore d'évaluation
- Resume-De-La-Physiologie-Digestive KHABET MOHAMED RAMZIDocument31 pagesResume-De-La-Physiologie-Digestive KHABET MOHAMED RAMZILachgar SalmaPas encore d'évaluation
- Conduite À Tenir Devant Une Sécheresse BuccaleDocument17 pagesConduite À Tenir Devant Une Sécheresse Buccaleammar eidPas encore d'évaluation
- SalivationDocument29 pagesSalivationAna MariaPas encore d'évaluation
- Motricité Oesophagienne 2021Document34 pagesMotricité Oesophagienne 2021Bek GhizlanePas encore d'évaluation
- Bonaz Bruno p01Document22 pagesBonaz Bruno p01NsangwaPas encore d'évaluation
- 01physio2an Resume-DigestiveDocument36 pages01physio2an Resume-DigestiveGlobule RougePas encore d'évaluation
- La Physiologie de La Digestion DR GuedjatiDocument23 pagesLa Physiologie de La Digestion DR GuedjatiMedMan686% (7)
- Chap 2 DigestionDocument10 pagesChap 2 DigestionSaloua ElboustatiPas encore d'évaluation
- Leaky Gut Syndrome pour les débutants Le livre d'entraide Comment interpréter correctement les symptômes d'un intestin perméable, en identifier les causes et soigner votre intestin étape par étapeD'EverandLeaky Gut Syndrome pour les débutants Le livre d'entraide Comment interpréter correctement les symptômes d'un intestin perméable, en identifier les causes et soigner votre intestin étape par étapePas encore d'évaluation
- 2023 06 26 BROCHURE ARCD FinaleDocument52 pages2023 06 26 BROCHURE ARCD FinaleSaad OuakiaaPas encore d'évaluation
- La Réplication d'ADN-Biologie Moléculaire-DahmaniDocument102 pagesLa Réplication d'ADN-Biologie Moléculaire-DahmaniSaad OuakiaaPas encore d'évaluation
- Cahiers de Le Ducation Et de La Formation N12 FRDocument243 pagesCahiers de Le Ducation Et de La Formation N12 FRSaad OuakiaaPas encore d'évaluation
- Corrigé RS 1è Session Bac Sénégal 2012 - Série S1Document1 pageCorrigé RS 1è Session Bac Sénégal 2012 - Série S1Saad OuakiaaPas encore d'évaluation
- Oasis SVT - 09.07Document32 pagesOasis SVT - 09.07Saad OuakiaaPas encore d'évaluation
- Mouvements À Force CentraleDocument27 pagesMouvements À Force CentraleSaad OuakiaaPas encore d'évaluation
- TP Biologie Cellulaire 2023-24Document23 pagesTP Biologie Cellulaire 2023-24Riri EssalhiPas encore d'évaluation
- COURS APC Tle Litteraire SVTDocument14 pagesCOURS APC Tle Litteraire SVTDalchy Makosso100% (1)
- Microbiologie Generale SVT S4 2021-2022Document47 pagesMicrobiologie Generale SVT S4 2021-2022sidibealhassane42Pas encore d'évaluation
- Dup (01) EvaluationsDocument5 pagesDup (01) EvaluationsOumarou Djibo ZoulkiffiPas encore d'évaluation
- DS 6 599 2018-2019 24-38-26-115Document8 pagesDS 6 599 2018-2019 24-38-26-115CEG1 DASSA-ZOUMEPas encore d'évaluation
- 5 - Tissu OsseuxDocument11 pages5 - Tissu OsseuxMey ZPas encore d'évaluation
- Control 2 s2 TCSF - CopieDocument2 pagesControl 2 s2 TCSF - CopieKhalid OualiPas encore d'évaluation
- 2 Chap 2 STRUCTURE PHYSIOLOGIE ET CROISSANCE BACT2RIENNEDocument62 pages2 Chap 2 STRUCTURE PHYSIOLOGIE ET CROISSANCE BACT2RIENNEBonaventure Babinne100% (1)
- Tri Proteine 2014 15Document131 pagesTri Proteine 2014 15chaimaePas encore d'évaluation
- Chapitre1+2 BiologieDocument48 pagesChapitre1+2 BiologieMohamed OuanaimPas encore d'évaluation
- Cours Microbiologieraccourci 1e Üre Anne Üe2007Document104 pagesCours Microbiologieraccourci 1e Üre Anne Üe2007Jo WiartPas encore d'évaluation
- TM Antibiotésistance V.FDocument47 pagesTM Antibiotésistance V.FAdri051Pas encore d'évaluation
- SVT - Fiche 1 - Organisation Fonctionnelle Du VivantDocument8 pagesSVT - Fiche 1 - Organisation Fonctionnelle Du VivantDjibril TourePas encore d'évaluation
- Structure Des BactériesDocument9 pagesStructure Des BactérieskhouildiPas encore d'évaluation
- Anatomie de La Cellule BactérienneDocument5 pagesAnatomie de La Cellule BactériennemalikaPas encore d'évaluation
- Biologie de La PulpeDocument22 pagesBiologie de La PulpeTania RomanPas encore d'évaluation
- Introduction MycologieDocument3 pagesIntroduction Mycologiebendib yacinePas encore d'évaluation
- HyaloplasmeDocument25 pagesHyaloplasmeamina44Pas encore d'évaluation
- Transmettre Information Genetique ExercicesDocument4 pagesTransmettre Information Genetique Exercicesmaimounadiallomaimouna10Pas encore d'évaluation
- 1 Généralites Sur La CelluleDocument40 pages1 Généralites Sur La CelluleOumarou Konta100% (1)
- 129 Biologie Cellulaire Introduction Au Vivant CPCMDocument17 pages129 Biologie Cellulaire Introduction Au Vivant CPCMFleur D'anisPas encore d'évaluation
- 2 Amaglo Ghana Prod Feuille Moringa2006Document132 pages2 Amaglo Ghana Prod Feuille Moringa2006Younes MakPas encore d'évaluation
- Bonnardel Jean Patrick - L'Argent ColloidalDocument70 pagesBonnardel Jean Patrick - L'Argent ColloidalCarlos SanmartinPas encore d'évaluation
- Cours de SVT (1S2)Document76 pagesCours de SVT (1S2)fallrosambayangPas encore d'évaluation
- 1ac AdilDocument2 pages1ac AdilAdil Adil0% (1)
- L3 BM Systématique Partie I PDFDocument18 pagesL3 BM Systématique Partie I PDFDjamila HEZILPas encore d'évaluation
- Risque de La Biologie MoléculaireDocument35 pagesRisque de La Biologie Moléculaireمولاي أمحمد عكرميPas encore d'évaluation
- TD Le Cytosquelette PolycopeDocument5 pagesTD Le Cytosquelette Polycopekirito 83Pas encore d'évaluation
- Bios 202 CoursDocument49 pagesBios 202 CoursMassaou Blama RapataPas encore d'évaluation