Vous aimerez peut-être aussi
- Laventure Ambigue - Laventure AmbigueDocument180 pagesLaventure Ambigue - Laventure AmbigueLavinia Checiu100% (4)
- Neurobiologie: Les Grands Articles d'UniversalisD'EverandNeurobiologie: Les Grands Articles d'UniversalisPas encore d'évaluation
- Guide Excel 2013Document27 pagesGuide Excel 2013Aliou Labayka100% (1)
- Muscles: Les Grands Articles d'UniversalisD'EverandMuscles: Les Grands Articles d'UniversalisPas encore d'évaluation
- Cours Biologie Cellulaire PDFDocument90 pagesCours Biologie Cellulaire PDFZį Zoū100% (1)
- RIMAH - Préambule Et IntroductionDocument30 pagesRIMAH - Préambule Et IntroductionDaaray Cheikhoul Xadim100% (26)
- Config Depannage PCDocument33 pagesConfig Depannage PCAliou LabaykaPas encore d'évaluation
- ReplicationDocument21 pagesReplicationMohamed Yassine MtarPas encore d'évaluation
- Embryologie GénéraleDocument44 pagesEmbryologie GénéraleMariem Nejima100% (1)
- QCM Corrigé en Embryologie 2004-2005 - Coursdemedecine - Free-HDocument14 pagesQCM Corrigé en Embryologie 2004-2005 - Coursdemedecine - Free-Hdeadbysunrisee100% (13)
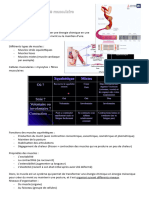
- Les Tissus MusculairesDocument56 pagesLes Tissus MusculairesIdrissou Fmsb100% (2)
- Exercices - Sujets de Cryptogamie-1Document11 pagesExercices - Sujets de Cryptogamie-1Aliou LabaykaPas encore d'évaluation
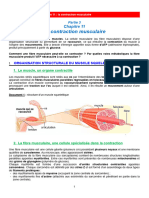
- La Contraction Musculaire PDFDocument9 pagesLa Contraction Musculaire PDFFaculté De Médecine Béchar50% (2)
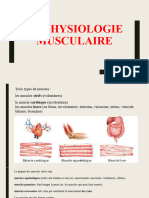
- Physiologie Musculaire 2020Document47 pagesPhysiologie Musculaire 2020drahlemtrifPas encore d'évaluation
- Fiches de Revision SP 1e SemestreDocument25 pagesFiches de Revision SP 1e SemestreAourras NourdinPas encore d'évaluation
- Contraction Musculaire CouleurDocument286 pagesContraction Musculaire Couleurmatis47215100% (1)
- La Contraction MusculaireDocument11 pagesLa Contraction MusculaireJihadi AbbouPas encore d'évaluation
- Surrénales: Les Grands Articles d'UniversalisD'EverandSurrénales: Les Grands Articles d'UniversalisPas encore d'évaluation
- Tissus MusculairesDocument33 pagesTissus MusculairessafemindPas encore d'évaluation
- Labo BébéDocument4 pagesLabo BébésalimPas encore d'évaluation
- Activité MsculaireDocument7 pagesActivité MsculaireameniPas encore d'évaluation
- NEUROPHYSIOLOGIEDocument47 pagesNEUROPHYSIOLOGIEPrince AlegaPas encore d'évaluation
- 2-Physiologie MusculaireDocument44 pages2-Physiologie MusculaireIslęm OuaribPas encore d'évaluation
- Physiologie Muscles Striés SquelettiquesDocument16 pagesPhysiologie Muscles Striés SquelettiquessafemindPas encore d'évaluation
- Physio1an31-7jonction NeuromusculaireDocument15 pagesPhysio1an31-7jonction Neuromusculaireelidrissimaryam733Pas encore d'évaluation
- Le Muscle NormalDocument41 pagesLe Muscle NormalHelena KenguePas encore d'évaluation
- CM Muscle FRB 6 DiaDocument13 pagesCM Muscle FRB 6 DiaWilly Le DruillennecPas encore d'évaluation
- P Motricité (C. Quairiaux) - 26.03.21Document5 pagesP Motricité (C. Quairiaux) - 26.03.21nasten16fhPas encore d'évaluation
- Les Tissus MusculairesDocument11 pagesLes Tissus MusculairesZoulkiffiPas encore d'évaluation
- Chapitre4.1 Biochimie Cellulaire Et FonctionnelleDocument7 pagesChapitre4.1 Biochimie Cellulaire Et Fonctionnelledraboissouf174Pas encore d'évaluation
- Physiologie Du Muscle Et de La Contraction MusculaireDocument5 pagesPhysiologie Du Muscle Et de La Contraction MusculaireImane BenderazPas encore d'évaluation
- 4.contraction MusculaireDocument40 pages4.contraction MusculaireKima MadPas encore d'évaluation
- ch10 Solutionnaire 1569538426Document12 pagesch10 Solutionnaire 1569538426Wilfried KouaoPas encore d'évaluation
- Les Muscles LissesDocument5 pagesLes Muscles LissesHassanElhassanPas encore d'évaluation
- La Contraction MusculaireDocument6 pagesLa Contraction MusculaireLim BunkimPas encore d'évaluation
- Le Fonctionnement Du Muscle Squelettique: Magazine de Sciences de La Vie Et de La TerreDocument28 pagesLe Fonctionnement Du Muscle Squelettique: Magazine de Sciences de La Vie Et de La TerreSifddin GuidPas encore d'évaluation
- Hand Physiologie MuscleDocument9 pagesHand Physiologie MuscleRachid BenPas encore d'évaluation
- tp7 CorrigeDocument4 pagestp7 CorrigeEmmanuel KoffiPas encore d'évaluation
- Support de Cours Physiologie Et Physiopathologie de La Cellule Musculaire ABED NousseibaDocument33 pagesSupport de Cours Physiologie Et Physiopathologie de La Cellule Musculaire ABED NousseibaSamwil SUANONPas encore d'évaluation
- 03 - 2 - Proprietes Mecaniques Du Muscle - ContractionDocument30 pages03 - 2 - Proprietes Mecaniques Du Muscle - ContractionGloria KitonPas encore d'évaluation
- CM Physiologie MusculaireDocument20 pagesCM Physiologie Musculairekouirass.mohamedPas encore d'évaluation
- 1.physiologie Musculaire 2023Document59 pages1.physiologie Musculaire 2023aminezebar3Pas encore d'évaluation
- MOTRICITE Par 2 DiapDocument79 pagesMOTRICITE Par 2 DiapRaphaël BikoahPas encore d'évaluation
- Tissus MusculairesDocument16 pagesTissus MusculairesandriePas encore d'évaluation
- Khaiter Devoir Numero 1Document5 pagesKhaiter Devoir Numero 1Y2kMagiikPas encore d'évaluation
- Muscles IFSI MZ 2027 Sept 2017 n37Document37 pagesMuscles IFSI MZ 2027 Sept 2017 n37Phil BarnyPas encore d'évaluation
- Diapo Physio Hennock-2Document31 pagesDiapo Physio Hennock-2mossila BradleyPas encore d'évaluation
- Boutonnat Jean p01 PDFDocument61 pagesBoutonnat Jean p01 PDFMoncef PechaPas encore d'évaluation
- Bilan Du Chapitre 11 - La Contraction MusculaireDocument12 pagesBilan Du Chapitre 11 - La Contraction MusculaireYousra INDIAPas encore d'évaluation
- Tissu Musculaire PDFDocument58 pagesTissu Musculaire PDFayoubPas encore d'évaluation
- Physiologie Animale Cours 15Document168 pagesPhysiologie Animale Cours 15g69wwpfw5kPas encore d'évaluation
- Le Muscle SquelettiqueDocument4 pagesLe Muscle SquelettiqueGabriela TituPas encore d'évaluation
- Physiologie Du MuscleDocument33 pagesPhysiologie Du MuscleYannick NdriPas encore d'évaluation
- Cours Unité 1 Chapitre 2 EL BoukhariDocument10 pagesCours Unité 1 Chapitre 2 EL BoukhariImene ImenePas encore d'évaluation
- Physiologie LA JONCTION NEUROMUSCULAIREDocument9 pagesPhysiologie LA JONCTION NEUROMUSCULAIREtoni.slmPas encore d'évaluation
- Physio Du MuscleDocument19 pagesPhysio Du Musclesamcruz31100% (1)
- Muscles Cours2Document3 pagesMuscles Cours2fardadi rachidPas encore d'évaluation
- Histo23 07-Tissu MusculaireDocument11 pagesHisto23 07-Tissu MusculaireSamwil SUANONPas encore d'évaluation
- La Physiologie MusculaireDocument51 pagesLa Physiologie MusculaireJoronavalona RASAMIMANANAPas encore d'évaluation
- 08-Physiologie de La Jonction NeuromusculaireDocument26 pages08-Physiologie de La Jonction NeuromusculaireBook HunterPas encore d'évaluation
- Physiologie Du Mucle Cardiaque 2Document34 pagesPhysiologie Du Mucle Cardiaque 2Marcus MilitoPas encore d'évaluation
- 7 - Les Tissus MusculairesDocument6 pages7 - Les Tissus MusculairesHayfa Ben taherPas encore d'évaluation
- Cours Mucle RénovéDocument11 pagesCours Mucle Rénovémohamed.aziz.amri.11Pas encore d'évaluation
- 21 Tspe M1 FicheDocument2 pages21 Tspe M1 FicheMathilde JaeckelPas encore d'évaluation
- Résumé U-1 - P-2Document4 pagesRésumé U-1 - P-2ygtzdntp2j100% (1)
- Chapitre 3 Le Système NerveuxDocument26 pagesChapitre 3 Le Système NerveuxdfhohfeicxhPas encore d'évaluation
- Physiologie Du Muscle Strie SquelettiqueDocument39 pagesPhysiologie Du Muscle Strie SquelettiqueRaouf ChikhiPas encore d'évaluation
- MOSTA Physiologie de La Jonction NeuromusculaireDocument27 pagesMOSTA Physiologie de La Jonction NeuromusculaireSamia BelabbasPas encore d'évaluation
- 14 Histophysiologie Du Tissu NerveuxDocument7 pages14 Histophysiologie Du Tissu NerveuxGeorge BorsPas encore d'évaluation
- La Plaque MotriceDocument22 pagesLa Plaque MotriceTourgout Alb CengizPas encore d'évaluation
- BIO 111 CHAP IXb 2021Document12 pagesBIO 111 CHAP IXb 2021meteweflorindaPas encore d'évaluation
- Control 2 s2 TCSF - CopieDocument2 pagesControl 2 s2 TCSF - CopieKhalid OualiPas encore d'évaluation
- Referentiel NACRES-juin2015Document127 pagesReferentiel NACRES-juin2015Abderraouf MouPas encore d'évaluation
- TDBR-P21 Corrige2Document17 pagesTDBR-P21 Corrige2Alejandro Domingo NúñezPas encore d'évaluation
- Compte Rendu Tp2 PCMDocument5 pagesCompte Rendu Tp2 PCMbahiamouchachou100% (1)
- PPT Les Épithéliums - PR AmegborDocument22 pagesPPT Les Épithéliums - PR AmegbormelchiPas encore d'évaluation
- Comment Le Muscle Strié Squelettique TransformeDocument19 pagesComment Le Muscle Strié Squelettique TransformeBentouina mohammed saidPas encore d'évaluation
- Nationaux SVTDocument113 pagesNationaux SVTTr LwPas encore d'évaluation
- Biologie Générale HumaineDocument10 pagesBiologie Générale HumaineSarah CabieuPas encore d'évaluation
- Production Et Role ATPDocument3 pagesProduction Et Role ATPWACDPas encore d'évaluation
- Cours 4 - Fécondation - Format 1Document15 pagesCours 4 - Fécondation - Format 1Nãřímäñ KŁayPas encore d'évaluation
- 0-Plan Genetique M2 1920Document3 pages0-Plan Genetique M2 1920Zahra YasminePas encore d'évaluation
- Acides Amines Et Proteines Biochimie Structurale-1Document104 pagesAcides Amines Et Proteines Biochimie Structurale-1Cheick SANOUPas encore d'évaluation
- Cytokines ChimiokinesDocument2 pagesCytokines Chimiokines9yky45jv7cPas encore d'évaluation
- QCM Immunologie Sans CorrectionDocument233 pagesQCM Immunologie Sans Correctionsalma chelliouiPas encore d'évaluation
- Bac SVT 2022 SVT-FR NorDocument6 pagesBac SVT 2022 SVT-FR Nor꧁TAKAYUKIPas encore d'évaluation
- 5-Pancréas Endocrine PDFDocument31 pages5-Pancréas Endocrine PDFthierno amadou bah100% (1)
- Azzedine ZouatiniDocument20 pagesAzzedine Zouatinialex mondomobePas encore d'évaluation
- Je Teste Mes Connaissances: 10 PTS: - Retrouvez Les Mots Correspondant Aux Définitions SuivantesDocument2 pagesJe Teste Mes Connaissances: 10 PTS: - Retrouvez Les Mots Correspondant Aux Définitions SuivantesGénéral Engongha MinangPas encore d'évaluation
- 2016 Cellules Et Tissus PDFDocument76 pages2016 Cellules Et Tissus PDFTita Tita FatimaPas encore d'évaluation
- 24-Regulation de La GlycemieDocument6 pages24-Regulation de La GlycemieSerigne Sohibou GayePas encore d'évaluation
- Fonctions TubulairesDocument52 pagesFonctions TubulairesLina Ray Ben100% (1)
- Devoir de Contrôle N°1 Avec Correction - SVT - 3ème Sciences Exp (2023-2024) MR HAOUARI ALIDocument6 pagesDevoir de Contrôle N°1 Avec Correction - SVT - 3ème Sciences Exp (2023-2024) MR HAOUARI ALInediasakliPas encore d'évaluation
- 3 L'électrophysiologie CardiaqueDocument8 pages3 L'électrophysiologie Cardiaqueaya fifiPas encore d'évaluation
- Interessant++++++++++++++Plan de Votre TheseDocument325 pagesInteressant++++++++++++++Plan de Votre Thesea.itatahinePas encore d'évaluation