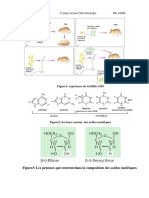
Vous aimerez peut-être aussi
- Biologie Moléculaire Chapitre 5 M1 1Document5 pagesBiologie Moléculaire Chapitre 5 M1 1اميرة في عرش ابيPas encore d'évaluation
- TraductionDocument36 pagesTraductionnonamethe427Pas encore d'évaluation
- Traduction L2 SADocument33 pagesTraduction L2 SAiliazPas encore d'évaluation
- Chapitre 5 2020Document27 pagesChapitre 5 2020Fayol MpayPas encore d'évaluation
- Chapitre 12 Synthèse Des Protéines (Transcription-Traduction)Document5 pagesChapitre 12 Synthèse Des Protéines (Transcription-Traduction)One Love LifePas encore d'évaluation
- Traduction 05 - 12 - 2023.Document43 pagesTraduction 05 - 12 - 2023.s.lakhloufi5682Pas encore d'évaluation
- Les Ribosomes Et La Synthèse Protéique B6Document10 pagesLes Ribosomes Et La Synthèse Protéique B6kim namjoonPas encore d'évaluation
- HAV504B - Chapitre 2Document24 pagesHAV504B - Chapitre 2Ylona RamondPas encore d'évaluation
- Cours 8 - La Synthèse ProtéiqueDocument8 pagesCours 8 - La Synthèse ProtéiqueFarouk Ben YahiaPas encore d'évaluation
- GENETIQUE FONDAMENTALE - Traduction - MoodleDocument5 pagesGENETIQUE FONDAMENTALE - Traduction - MoodleGh7Pas encore d'évaluation
- La Transcription l3 Microbiologie 2022Document8 pagesLa Transcription l3 Microbiologie 2022rou röuPas encore d'évaluation
- La Synthèse ProtéiqueDocument6 pagesLa Synthèse ProtéiquealiPas encore d'évaluation
- GENETIQUE FONDAMENTALE - Transcription - MoodleDocument3 pagesGENETIQUE FONDAMENTALE - Transcription - MoodleGh7Pas encore d'évaluation
- Epissage Chez Les EucaryotesDocument6 pagesEpissage Chez Les EucaryotesAlain Bertrand JuniorPas encore d'évaluation
- CM04 - Traduction Et Routage Des ProteinesDocument8 pagesCM04 - Traduction Et Routage Des ProteinesRock In BandPas encore d'évaluation
- Synthèses Des ProtéinesDocument1 pageSynthèses Des ProtéinesAl sarahPas encore d'évaluation
- Exe171 SynthDocument6 pagesExe171 SynthModou LôPas encore d'évaluation
- Exe171 SynthDocument6 pagesExe171 SynthMahmoud WattPas encore d'évaluation
- La TraductionDocument2 pagesLa Traductionnadirbelhadj39Pas encore d'évaluation
- Activité 7 Traduction Chapitre 1 Unité 2Document3 pagesActivité 7 Traduction Chapitre 1 Unité 2hamamielloPas encore d'évaluation
- Cours 4 - DIAPO - ADN Aux ProtéinesDocument40 pagesCours 4 - DIAPO - ADN Aux Protéinesbyy.monkey emzPas encore d'évaluation
- Comparaison de L'expression Génique Chez Les Eucaryotes Et Chez Les BactériesDocument3 pagesComparaison de L'expression Génique Chez Les Eucaryotes Et Chez Les BactériesNess LzPas encore d'évaluation
- SynthèseDocument10 pagesSynthèseElvy prices Sampinou mpompaPas encore d'évaluation
- 3 Synthèse Protéique-2Document12 pages3 Synthèse Protéique-2Sagacious IvejutenPas encore d'évaluation
- Genetique1an Transcription Adn2017hanachiDocument37 pagesGenetique1an Transcription Adn2017hanachisoumeya soltaniPas encore d'évaluation
- 9 Transcription Chap IV Expression Inf GenDocument9 pages9 Transcription Chap IV Expression Inf GenRock In BandPas encore d'évaluation
- CHEIDIDocument4 pagesCHEIDIcheilla nandjioPas encore d'évaluation
- BG3 ExosDocument22 pagesBG3 Exospersonne12Pas encore d'évaluation
- 16-Traduction de L'information GénétiqueDocument33 pages16-Traduction de L'information GénétiqueInsafPas encore d'évaluation
- 1 TranscriptionDocument12 pages1 TranscriptionYounes YouyouPas encore d'évaluation
- BMGG La TranscriptionDocument22 pagesBMGG La TranscriptionAbbou BelkaisPas encore d'évaluation
- Cours BMGG 2018Document43 pagesCours BMGG 2018Fares ManelPas encore d'évaluation
- BCM 7 Transcription TraductionDocument56 pagesBCM 7 Transcription TraductionJoel AmaniPas encore d'évaluation
- Biologie Moléculaire Et GG 2Document30 pagesBiologie Moléculaire Et GG 2Sangaré soungaloPas encore d'évaluation
- SVT TA - L7 - La Biosynthèse Des ProtéinesDocument12 pagesSVT TA - L7 - La Biosynthèse Des Protéinestangaramariam566Pas encore d'évaluation
- Exercices D'application Chapitre 3 - La Synthèse Des Protéines.Document2 pagesExercices D'application Chapitre 3 - La Synthèse Des Protéines.radietbrahim21Pas encore d'évaluation
- Ed Biomol 2Document2 pagesEd Biomol 2Alexia HritcuPas encore d'évaluation
- Chapitre Syntha Se ProtaiqueDocument29 pagesChapitre Syntha Se ProtaiqueNouha HadjPas encore d'évaluation
- Système Endomembranaire - NotesDocument18 pagesSystème Endomembranaire - Notesemiliedurosoir2Pas encore d'évaluation
- 3eme Biochimie 2019-2020 Mahdab N L3 Bioch COURS DE BIO MOLDocument22 pages3eme Biochimie 2019-2020 Mahdab N L3 Bioch COURS DE BIO MOLry yukPas encore d'évaluation
- Début Chapitre 3 Expression Du Patrimoine Génétique 2023-2024Document3 pagesDébut Chapitre 3 Expression Du Patrimoine Génétique 2023-2024cloe.garrotPas encore d'évaluation
- SVT 1ère D - L5 - La Synthèse Des ProtéinesDocument12 pagesSVT 1ère D - L5 - La Synthèse Des ProtéinesYedson GrâcePas encore d'évaluation
- CHAPITRE 5 Expression Du Patrimoine GenetiqueDocument10 pagesCHAPITRE 5 Expression Du Patrimoine GenetiqueVictor BillardPas encore d'évaluation
- ! Code Genetique. Biosynthese Des Proteines Et AminoacidesDocument27 pages! Code Genetique. Biosynthese Des Proteines Et AminoacidesMarii AlionaPas encore d'évaluation
- Le GèneDocument4 pagesLe GèneRaian SabahPas encore d'évaluation
- Evaluation 2Document5 pagesEvaluation 2firdaoussaoud22Pas encore d'évaluation
- Génétique 6Document2 pagesGénétique 6hkuuiPas encore d'évaluation
- 14.03.2016 La TraductionDocument11 pages14.03.2016 La TraductionEaPas encore d'évaluation
- Cours 2 MTDocument65 pagesCours 2 MTحسن غالية حسن غاليةPas encore d'évaluation
- Chapitre I: L'expression de L'information GénétiqueDocument74 pagesChapitre I: L'expression de L'information Génétiqueahméd_facebPas encore d'évaluation
- BIOLOGIE MOLÉCULAIRE - Expression Des GènesDocument16 pagesBIOLOGIE MOLÉCULAIRE - Expression Des GènesVIALEPas encore d'évaluation
- BiochDocument243 pagesBiochyouness.khalfaouiPas encore d'évaluation
- Cours Transcription L2SADocument16 pagesCours Transcription L2SAiliazPas encore d'évaluation
- TRAnscriptionDocument16 pagesTRAnscriptionmaysounPas encore d'évaluation
- Cours 5 La TranscriptionDocument13 pagesCours 5 La Transcriptiondoukarafousseyni7Pas encore d'évaluation
- Med Genetique-Transcription EucaryotesDocument20 pagesMed Genetique-Transcription EucaryotesRacha MalakPas encore d'évaluation
- Exposé Du Groupe 9Document35 pagesExposé Du Groupe 9Grâce KABOREPas encore d'évaluation
- A Synthèse Des ProtéinesDocument3 pagesA Synthèse Des ProtéinesNadaPas encore d'évaluation
- Correction de La Serie IVDocument3 pagesCorrection de La Serie IVakboub.ilyasPas encore d'évaluation
- 5 EstomacDocument6 pages5 Estomacsevda mouviePas encore d'évaluation
- Péritoine Mimo AmineDocument5 pagesPéritoine Mimo Aminelydouche0805Pas encore d'évaluation
- Sels Minéraux N.MDocument14 pagesSels Minéraux N.Mlydouche0805Pas encore d'évaluation
- 5-Digestion Et Absorption IntestinaleDocument3 pages5-Digestion Et Absorption Intestinalesevda mouviePas encore d'évaluation
- Guide Ayoub Yaici (2015-2016)Document4 pagesGuide Ayoub Yaici (2015-2016)Just KidnessPas encore d'évaluation
- 3-Physiologie GastriqueDocument5 pages3-Physiologie Gastriquesevda mouviePas encore d'évaluation
- 13-Outils de Biologie Moléculaire 1 N.MDocument5 pages13-Outils de Biologie Moléculaire 1 N.Msevda mouviePas encore d'évaluation
- 10-Modifications Post Traductionnelle N.MDocument3 pages10-Modifications Post Traductionnelle N.Msevda mouviePas encore d'évaluation
- Communication CellulaireDocument4 pagesCommunication Cellulairesevda mouviePas encore d'évaluation
- 1-Applications de La Génétique Dans Le Domaine Médical N.MDocument5 pages1-Applications de La Génétique Dans Le Domaine Médical N.MAmina AmiinaPas encore d'évaluation
- Métabolisme Des LipidesDocument7 pagesMétabolisme Des Lipidessevda mouviePas encore d'évaluation
- Système Nerveux AutonomeDocument43 pagesSystème Nerveux AutonomeAbderrahmane SarPas encore d'évaluation
- Exercices Cours3Document6 pagesExercices Cours3SAMI DHAOUI100% (1)
- ThalophytesDocument40 pagesThalophytesAbderrahim KharroubiPas encore d'évaluation
- Chromosomes Et CaryotypeDocument19 pagesChromosomes Et CaryotypejhbPas encore d'évaluation
- BMC Fiches UE M1S1 - 2022-2023Document30 pagesBMC Fiches UE M1S1 - 2022-2023Khánh Lê VănPas encore d'évaluation
- Embryo OursinDocument91 pagesEmbryo Oursintraore al-saphirPas encore d'évaluation
- Fiche3 GentiqueDocument2 pagesFiche3 GentiquecarlotagonztapPas encore d'évaluation
- Exe171 SynthDocument6 pagesExe171 SynthModou LôPas encore d'évaluation
- Anti Cyclines PDFDocument8 pagesAnti Cyclines PDFWassCANPas encore d'évaluation
- Fiche Pedagogique 6eme Fiche Pedagogique 6emeDocument106 pagesFiche Pedagogique 6eme Fiche Pedagogique 6ememonsieur web informaticienPas encore d'évaluation
- TD 3 Synthèse ProtéiqueDocument2 pagesTD 3 Synthèse ProtéiqueMohamed Badreddine Mokhtari100% (2)
- Exercice Polydactylie CorrectionDocument1 pageExercice Polydactylie Correctionnaisbais786Pas encore d'évaluation
- Chapitre Syntha Se ProtaiqueDocument29 pagesChapitre Syntha Se ProtaiqueNouha HadjPas encore d'évaluation
- Cours Génétique LPSVT S3Document87 pagesCours Génétique LPSVT S3samifilol703Pas encore d'évaluation
- GENETIQUE FONDAMENTALE - Transcription - MoodleDocument3 pagesGENETIQUE FONDAMENTALE - Transcription - MoodleGh7Pas encore d'évaluation
- Nature de L'information Génétique ExercicesDocument3 pagesNature de L'information Génétique Exercicesayaayasebaba1Pas encore d'évaluation
- Cycles Biologiques Et Structures Reproductrices Chap06Document24 pagesCycles Biologiques Et Structures Reproductrices Chap06almnaouarPas encore d'évaluation
- LA REPRODUCTION CHEZ L'HOMME Cours TotalDocument11 pagesLA REPRODUCTION CHEZ L'HOMME Cours TotalMohamed OUENJLI MATHBOUTPas encore d'évaluation
- La Reproduction Humaine Cours SVT 2ACDocument7 pagesLa Reproduction Humaine Cours SVT 2ACSalah Eddine BelmouffeqPas encore d'évaluation
- Bac Blanc 2020 SVT Serie DDocument8 pagesBac Blanc 2020 SVT Serie DDan LOUBINDOUPas encore d'évaluation
- Devoir - À - Remettre Modelisation Biologie MarineDocument4 pagesDevoir - À - Remettre Modelisation Biologie MarineRabha EnanehcPas encore d'évaluation
- Embryologie - DIAPO - Introduction + Semaine 1Document57 pagesEmbryologie - DIAPO - Introduction + Semaine 1surgeon forgeronPas encore d'évaluation
- Devoir de Contrôle N°2 - SVT - Bac Sciences Exp (2017-2018) Mme Mbarka Harbawi PDFDocument4 pagesDevoir de Contrôle N°2 - SVT - Bac Sciences Exp (2017-2018) Mme Mbarka Harbawi PDFMohamed SaidiPas encore d'évaluation
- Biologie Végétale CHAPITRE 5Document7 pagesBiologie Végétale CHAPITRE 5MįSs BęãüTÿPas encore d'évaluation
- UE2 - Biologie Cellulaire - Production D - Énergie Dans La CelluleDocument16 pagesUE2 - Biologie Cellulaire - Production D - Énergie Dans La Cellulesun-nee-chan9Pas encore d'évaluation
- Activité 1 Cycle CellulaireDocument4 pagesActivité 1 Cycle CellulaireLeane GervasoniPas encore d'évaluation
- Embryologie.2020-2021.partie 2 (Mode de Compatibilité)Document19 pagesEmbryologie.2020-2021.partie 2 (Mode de Compatibilité)rekik hibaPas encore d'évaluation
- Med Genetique-Transcription ProcaryotesDocument17 pagesMed Genetique-Transcription ProcaryotesRacha MalakPas encore d'évaluation
- DD-Régulation de L'expression Des Gènes EucaryotesDocument3 pagesDD-Régulation de L'expression Des Gènes Eucaryotesdora_stellaPas encore d'évaluation
- I Modification Post TranscriptionnelleDocument25 pagesI Modification Post TranscriptionnelleFarida SirimaPas encore d'évaluation
- SujetL2BCGS 2013 Semestre2Document1 pageSujetL2BCGS 2013 Semestre2Cheikh ThiamPas encore d'évaluation