Vous aimerez peut-être aussi
- Le Cycle Cellulaire: Données Biologiques Et Thérapies Ciblant Les cyclines/CDKDocument7 pagesLe Cycle Cellulaire: Données Biologiques Et Thérapies Ciblant Les cyclines/CDKfy fyPas encore d'évaluation
- Mecanismes Moleculaires Controlant Le CyDocument21 pagesMecanismes Moleculaires Controlant Le CymohamedPas encore d'évaluation
- Medsci 2003192173Document15 pagesMedsci 2003192173mohamedPas encore d'évaluation
- Corticoïdes Et Choc SeptiqueDocument10 pagesCorticoïdes Et Choc SeptiqueBENCHABANE OUSSAMAPas encore d'évaluation
- Antigène CarcinoembryonnaireDocument5 pagesAntigène CarcinoembryonnaireCDM achiffaPas encore d'évaluation
- TD Signal 2023Document10 pagesTD Signal 2023salym.mourid84Pas encore d'évaluation
- Cycle Cellulaire RESUMEDocument27 pagesCycle Cellulaire RESUMEmounia.34Pas encore d'évaluation
- A Thérapie Génique Du Cancer: Désir Mythique Ou Réalité Thérapeutique de Demain ?Document5 pagesA Thérapie Génique Du Cancer: Désir Mythique Ou Réalité Thérapeutique de Demain ?Hajar BoughroudPas encore d'évaluation
- Thérapies Ciblées 231010 165853Document5 pagesThérapies Ciblées 231010 165853moussaoued493Pas encore d'évaluation
- Cycle CellulaireDocument29 pagesCycle CellulairetinaPas encore d'évaluation
- Analyse Partie Disscussion (Finale)Document5 pagesAnalyse Partie Disscussion (Finale)kaouthar boumaichePas encore d'évaluation
- I07. Cytokines (Cour)Document7 pagesI07. Cytokines (Cour)Hibatallah NamounePas encore d'évaluation
- 2 NDDocument11 pages2 NDsamy.cheh18Pas encore d'évaluation
- TlouDocument26 pagesTlouChimie CleanPas encore d'évaluation
- Colchicine Dans La Goutte Usage Et MésusageDocument6 pagesColchicine Dans La Goutte Usage Et MésusageAmine DounanePas encore d'évaluation
- 2 - Enzymes Sériques Et TissulairesDocument19 pages2 - Enzymes Sériques Et TissulairesAbdou Ess50% (2)
- Mucovisidose Article 3Document5 pagesMucovisidose Article 3riyad saksakPas encore d'évaluation
- Medsci 2012285 P 465Document4 pagesMedsci 2012285 P 465KougangPas encore d'évaluation
- Aurora KinaseDocument9 pagesAurora KinasealainastierPas encore d'évaluation
- 15Document6 pages15lile aichaPas encore d'évaluation
- 23 Guimbaud 2 619 v2Document9 pages23 Guimbaud 2 619 v2Moncef CherifPas encore d'évaluation
- Les Cytokines 2019 2020Document7 pagesLes Cytokines 2019 2020tara mouradPas encore d'évaluation
- Survie Cellulaire Cancers MecanismesDocument7 pagesSurvie Cellulaire Cancers Mecanismestaoufik akabliPas encore d'évaluation
- 6-Cytotoxicité Nécrose ApoptoseDr - Boudemagh KDocument11 pages6-Cytotoxicité Nécrose ApoptoseDr - Boudemagh KAnouar BezzichePas encore d'évaluation
- 315 022 FotsingDocument10 pages315 022 FotsingBamory OuattaraPas encore d'évaluation
- Valeurs Usuelles de La Composition Du Liquide Céphalorachidien Chez Le Nouveau-NéDocument4 pagesValeurs Usuelles de La Composition Du Liquide Céphalorachidien Chez Le Nouveau-Néمحمد رؤوف خير الدينPas encore d'évaluation
- CHIMIOTHERAPIEDocument3 pagesCHIMIOTHERAPIEcopie master100% (1)
- Mécanismes D'action Des Inhibiteurs de Kinases Cyclines-Dépendantes: Identification de Leurs Cibles Intracellulaires Par Chromatographie D'affinitéDocument162 pagesMécanismes D'action Des Inhibiteurs de Kinases Cyclines-Dépendantes: Identification de Leurs Cibles Intracellulaires Par Chromatographie D'affinitéIsocrate DourrawaPas encore d'évaluation
- 15-Phelip Jean-Marc Clavel Lea Et Rinaldi Leslie-TexteDocument8 pages15-Phelip Jean-Marc Clavel Lea Et Rinaldi Leslie-TexteLylou AmayouPas encore d'évaluation
- 10 - Cycle Et Senescence Diff Et Mort Cellulaire Cours 2022Document92 pages10 - Cycle Et Senescence Diff Et Mort Cellulaire Cours 2022Thuy TranPas encore d'évaluation
- Physiologie de La Contraction CardiaqueDocument10 pagesPhysiologie de La Contraction CardiaqueHassan OulehriPas encore d'évaluation
- Cours 1Document19 pagesCours 1Kimberley MétivierPas encore d'évaluation
- SdarticleDocument8 pagesSdarticleha.boussoussaPas encore d'évaluation
- Les Espèces Actives de L'oxygène: Le Yin Et Le Yang de La MitochondrieDocument7 pagesLes Espèces Actives de L'oxygène: Le Yin Et Le Yang de La MitochondrieZaki NaniPas encore d'évaluation
- Récepteurs Activables Par Les Proliférateurs Des Peroxysomes (PPAR) : Leur Implication Dans Le Métabolisme Des Hydrates de Carbone Et Des LipidesDocument9 pagesRécepteurs Activables Par Les Proliférateurs Des Peroxysomes (PPAR) : Leur Implication Dans Le Métabolisme Des Hydrates de Carbone Et Des LipidesshafikPas encore d'évaluation
- Biologie 2016Document15 pagesBiologie 2016Mohbio MohaPas encore d'évaluation
- Plasma Membrane Calcium Pumps in SmoothDocument12 pagesPlasma Membrane Calcium Pumps in SmoothSergio Del AngelPas encore d'évaluation
- Bac S 2018 - Amérique Du Sud - SVT - Académie de BesançonDocument10 pagesBac S 2018 - Amérique Du Sud - SVT - Académie de Besançongzqg9cp7mtPas encore d'évaluation
- Les Glucocorticoides 27 Février 2020 À ImprimerDocument40 pagesLes Glucocorticoides 27 Février 2020 À ImprimerGnaly KpazaiPas encore d'évaluation
- Marqueurs Cardiaques - DES Déc2022 (Mode de Compatibilité) - PR Gobeaux - COCHINDocument54 pagesMarqueurs Cardiaques - DES Déc2022 (Mode de Compatibilité) - PR Gobeaux - COCHINNour AngriniPas encore d'évaluation
- 5-Cycle ÇDocument25 pages5-Cycle ÇMohamed OuHouissPas encore d'évaluation
- Poster GenetiqueDocument1 pagePoster GenetiqueRanine SaadaouiPas encore d'évaluation
- Cycle Partie2 1diaDocument32 pagesCycle Partie2 1diaAliBenAhmedPas encore d'évaluation
- Fiche II TD BIO 111 2020-2021Document2 pagesFiche II TD BIO 111 2020-2021Pius NdjebetPas encore d'évaluation
- Cellule Cancéreuse Et BioinformatiqueDocument4 pagesCellule Cancéreuse Et Bioinformatiquepanopoulos_295063483Pas encore d'évaluation
- CytotoxicitéDocument7 pagesCytotoxicitéAl KhadaffyPas encore d'évaluation
- C M 2-Oncologie I-2016Document111 pagesC M 2-Oncologie I-2016NanfaPas encore d'évaluation
- Le Rôle Du Noyau Du TractusDocument4 pagesLe Rôle Du Noyau Du Tractusmilenerato2240Pas encore d'évaluation
- Bac SVT 2Document8 pagesBac SVT 2COLOMBIPas encore d'évaluation
- Retard Mitotique Et Maintien de L'intégrité ChromosomiqueDocument28 pagesRetard Mitotique Et Maintien de L'intégrité ChromosomiqueAnis Wins50% (2)
- Les Anticorps Monoclonaux en Hematologie en 2009Document7 pagesLes Anticorps Monoclonaux en Hematologie en 2009ruthdjeketPas encore d'évaluation
- He Et Al 2014 - HCNDocument121 pagesHe Et Al 2014 - HCNbruno paivaPas encore d'évaluation
- TD 1 Biologie Cellulaire Et Moleculaire de Developpement m1 Ba1Document22 pagesTD 1 Biologie Cellulaire Et Moleculaire de Developpement m1 Ba1Ždn ÂśšiaPas encore d'évaluation
- 4-Chimiothérapie 2023Document27 pages4-Chimiothérapie 2023Moulaye MedPas encore d'évaluation
- Introduction Pharmaco. Generale m1 s1 G.PDocument13 pagesIntroduction Pharmaco. Generale m1 s1 G.PHamdaoui dounia100% (1)
- 8 HomingDocument50 pages8 HomingFarah SalhiPas encore d'évaluation
- 9 - Cellules Cancéreuses Et Tissu Tumoral Déf ÉtudiantsDocument45 pages9 - Cellules Cancéreuses Et Tissu Tumoral Déf ÉtudiantsChamalix ChamalixPas encore d'évaluation
- Traitements Immunosuppresseurs MecanismeDocument16 pagesTraitements Immunosuppresseurs MecanismeAlain SoucotPas encore d'évaluation
- Tumeurs Neuro Endocrines 2017 PDFDocument28 pagesTumeurs Neuro Endocrines 2017 PDFWassCANPas encore d'évaluation
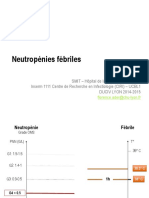
- 143 Neutropenies AgranulocytosesDocument8 pages143 Neutropenies AgranulocytosesWassCANPas encore d'évaluation
- Tumeurs Neuro Endocrines 2017 PDFDocument28 pagesTumeurs Neuro Endocrines 2017 PDFWassCANPas encore d'évaluation
- 2018 Gerrico NFDocument130 pages2018 Gerrico NFWassCANPas encore d'évaluation
- 2015 DUCIV Lyon Ader - Neutropenies - Febriles PDFDocument62 pages2015 DUCIV Lyon Ader - Neutropenies - Febriles PDFWassCANPas encore d'évaluation
- 2017-Neutropenies-Febriles-Alfandari (1) 2Document75 pages2017-Neutropenies-Febriles-Alfandari (1) 2WassCANPas encore d'évaluation
- 2017-Neutropenies-Febriles-Alfandari (1) 2Document75 pages2017-Neutropenies-Febriles-Alfandari (1) 2WassCANPas encore d'évaluation
- Fiche Aplasie Febrile Oncomip - Decembre2017Document2 pagesFiche Aplasie Febrile Oncomip - Decembre2017WassCANPas encore d'évaluation
- 2018 Gerrico NFDocument130 pages2018 Gerrico NFWassCANPas encore d'évaluation
- AFSOS 2018 - Focus Sur Les Recommandations 2018 Concernant La Neutropénie Associée Aux Traitements - UnivadisDocument3 pagesAFSOS 2018 - Focus Sur Les Recommandations 2018 Concernant La Neutropénie Associée Aux Traitements - UnivadisWassCANPas encore d'évaluation
- Effets Indesirables Cutaneo Muqueux Des ChimiotherapiesDocument6 pagesEffets Indesirables Cutaneo Muqueux Des ChimiotherapiesWassCANPas encore d'évaluation
- Febrile Neutropenia PreventionDocument118 pagesFebrile Neutropenia PreventionWassCANPas encore d'évaluation
- 143 Neutropenies AgranulocytosesDocument8 pages143 Neutropenies AgranulocytosesWassCANPas encore d'évaluation
- NeutropeniefebrileDocument1 pageNeutropeniefebrileWassCANPas encore d'évaluation
- 2014-12 Ref Soin Support Anemie Cancer Afsos Pro PDFDocument22 pages2014-12 Ref Soin Support Anemie Cancer Afsos Pro PDFWassCANPas encore d'évaluation
- 1 Tester BRCA Ou HRD Quand PDFDocument30 pages1 Tester BRCA Ou HRD Quand PDFWassCANPas encore d'évaluation
- L'Anemie Chimio-Induite - Chemotherapy-Induced AnemiaDocument141 pagesL'Anemie Chimio-Induite - Chemotherapy-Induced AnemiaWassCANPas encore d'évaluation
- Ase Cancerologie - Assurance Maladie PDFDocument8 pagesAse Cancerologie - Assurance Maladie PDFWassCANPas encore d'évaluation
- Agents Stimulant L'érythropoïèse Et Prise en Charge de L'anémie Chimio-Induite: Point D'actualitéDocument4 pagesAgents Stimulant L'érythropoïèse Et Prise en Charge de L'anémie Chimio-Induite: Point D'actualitéWassCANPas encore d'évaluation
- L'Hepcidine, La Grande Dame Du Fer: Institut Cochin, ParisDocument5 pagesL'Hepcidine, La Grande Dame Du Fer: Institut Cochin, ParisWassCANPas encore d'évaluation
- 1 Tester BRCA Ou HRD QuandDocument30 pages1 Tester BRCA Ou HRD QuandWassCANPas encore d'évaluation
- 6 Prise en Charge Tumeur Epitheliale Rare OvaireDocument17 pages6 Prise en Charge Tumeur Epitheliale Rare OvaireWassCANPas encore d'évaluation
- 4 Prise en Charge Tumeur Germinale OvarienneDocument25 pages4 Prise en Charge Tumeur Germinale OvarienneWassCANPas encore d'évaluation
- Abc-304319-Accumulation de La Proteine p53 Et Surexpression Du Recepteur Her3 Dans Le Cancer Colorectal - W8uYVn8AAQEAAGL4opUAAAAC-aDocument7 pagesAbc-304319-Accumulation de La Proteine p53 Et Surexpression Du Recepteur Her3 Dans Le Cancer Colorectal - W8uYVn8AAQEAAGL4opUAAAAC-aWassCANPas encore d'évaluation
- Pathologies ORL Généralités - I.BREUSKINDocument89 pagesPathologies ORL Généralités - I.BREUSKINWassCANPas encore d'évaluation
- Action Mondiale Contre Le CancerCoverDocument15 pagesAction Mondiale Contre Le CancerCoverWassCANPas encore d'évaluation
- Kinases Dépendantes Des Cyclines: Rôle Biologique Et Implications Dans La Pathologie HumaineDocument9 pagesKinases Dépendantes Des Cyclines: Rôle Biologique Et Implications Dans La Pathologie HumaineWassCANPas encore d'évaluation
- Etude Cavum Maroc 2018 PDFDocument7 pagesEtude Cavum Maroc 2018 PDFWassCANPas encore d'évaluation
- Classification TNMDocument5 pagesClassification TNMWassCANPas encore d'évaluation
- Noyau InterphasiqueDocument9 pagesNoyau InterphasiqueDear tynaPas encore d'évaluation
- COURS BC - PlasteDocument23 pagesCOURS BC - PlasteMohammed El MachmoulPas encore d'évaluation
- Annales de Biocell-QcmDocument6 pagesAnnales de Biocell-QcmmedPas encore d'évaluation
- Correction ED Métabolisme Des Lipides 2020 2021 1Document32 pagesCorrection ED Métabolisme Des Lipides 2020 2021 1Belkis SioudPas encore d'évaluation
- 3-Enzyme 21-22Document40 pages3-Enzyme 21-22varyvira6677Pas encore d'évaluation
- Chapitre 12 Synthèse Des Protéines (Transcription-Traduction)Document5 pagesChapitre 12 Synthèse Des Protéines (Transcription-Traduction)One Love LifePas encore d'évaluation
- CM Regulation Metabolique 2 Enzymes Et Allosterie 2019 ModDocument31 pagesCM Regulation Metabolique 2 Enzymes Et Allosterie 2019 ModSirina LadjPas encore d'évaluation
- Le Monde Complexe Et Mouvant Des ARN.Document22 pagesLe Monde Complexe Et Mouvant Des ARN.tawfiq bouyahiaPas encore d'évaluation
- Cycle de KrebsDocument37 pagesCycle de KrebscepPas encore d'évaluation
- Cours 7 - RibosomesDocument5 pagesCours 7 - Ribosomesamina44Pas encore d'évaluation
- Métabolisme Des LipidesDocument17 pagesMétabolisme Des LipidesMei Wan100% (1)
- Cycle de KrebsDocument1 pageCycle de KrebsHadrien De GreefPas encore d'évaluation
- La Reproduction Sexuée Et Les Étapes de La Méiose - Résumé Et Révision - SVT - SchoolMouv PDFDocument3 pagesLa Reproduction Sexuée Et Les Étapes de La Méiose - Résumé Et Révision - SVT - SchoolMouv PDFjuini rawiaPas encore d'évaluation
- QSM BiochimieDocument25 pagesQSM BiochimieLGGPRONEPas encore d'évaluation
- 2 - Corrigé TD Chapitre 3 AllostérieDocument3 pages2 - Corrigé TD Chapitre 3 AllostérieChaka TraorePas encore d'évaluation
- La Respiration Cellulaire 1 PDFDocument27 pagesLa Respiration Cellulaire 1 PDFyyyyy100% (1)
- TD RecapitulatifDocument4 pagesTD RecapitulatifZa Hra100% (2)
- !!!!!!!!cours Biologie Moleculaie S5Document46 pages!!!!!!!!cours Biologie Moleculaie S5Miro100% (1)
- Voir Le Cours Sur Les Cinétiques À 2 Substrats.: Enzymologie: Réactions Enzymatiques À Deux SubstratsDocument5 pagesVoir Le Cours Sur Les Cinétiques À 2 Substrats.: Enzymologie: Réactions Enzymatiques À Deux SubstratsLAMINE DEGUISTARPas encore d'évaluation
- UntitledtdugogrDocument3 pagesUntitledtdugogrLoubna MichaPas encore d'évaluation
- Devoir Harmonise Avec Correction N°2 PDDocument7 pagesDevoir Harmonise Avec Correction N°2 PDElie ZRAPas encore d'évaluation
- Copie2 TWODocument7 pagesCopie2 TWOKhalid Shorts0% (1)
- td3 GenetiqueDocument8 pagestd3 Genetiquedjassa djassaPas encore d'évaluation
- Série 2-Génétique 2020-2021 DendougaDocument3 pagesSérie 2-Génétique 2020-2021 DendougaDou Aae100% (1)
- Cinétique À Deux Substrat.mDocument9 pagesCinétique À Deux Substrat.mKEBIR ZAHRAPas encore d'évaluation
- ARN InterférenceDocument3 pagesARN InterférenceshafikPas encore d'évaluation
- Thérapies Moléculaires Ciblées Et Antiangiogéniques Dans Le Traitement Des GlioblastomesDocument13 pagesThérapies Moléculaires Ciblées Et Antiangiogéniques Dans Le Traitement Des GlioblastomesyaalaPas encore d'évaluation
- Examen L1 Biologie Moléculaire 2005 1Document9 pagesExamen L1 Biologie Moléculaire 2005 1R-winPas encore d'évaluation
- TD Biochimie Métabolique 2022 2023Document5 pagesTD Biochimie Métabolique 2022 2023AG PROMO MarocPas encore d'évaluation
- 10 (6) - Variations Génétiques Et Réparation de l'ADN 6Document7 pages10 (6) - Variations Génétiques Et Réparation de l'ADN 6SaibiPas encore d'évaluation